Gastropoda
 Gastropods, the group of mollusks that include the terrestrial snails and slugs and the marine limpets, periwinkles, abalones, whelks, and their relatives, are the largest and most varied class of mollusks, with more than 75,000 extant species in addition to 15,000 fossil forms that are known.
Gastropods, the group of mollusks that include the terrestrial snails and slugs and the marine limpets, periwinkles, abalones, whelks, and their relatives, are the largest and most varied class of mollusks, with more than 75,000 extant species in addition to 15,000 fossil forms that are known.
These successful animals are the only class of mollusks to occupy both marine, fresh-water, and terrestrial environments. In the sea, most species are epibenthic, crawling on the surface, but there are some swimming forms as well (some types of Opisthobranchia). Although many gastropods are herbivorous grazers, several groups are active carnivores able to drill through the shell of their victim, or in the case of predatory forms like the Conidae (Neogastropoda) cases swallow it whole through a proboscis.
Most of the gastropods are classified on the characteristics the gill structures and other soft-bodied features. Few distinguishing characters of the shell are used in classification as many are the result of convergent evolution. Although the differences in the shell form may be difficult to recognize, different morphologic groups can generally can be differentiated on characteristics of ornamentation, shell shape, and aperture. The shell of many gastropods can either be external or, less commonly, internal. The difference can often be deduced by the luster of shell material and the presence of other features such as deviations of a structural shell form.
Gastropods are also sometimes known in older books as "Univalves", because they possess a single shell (in contrast to bivalves with two, and chitons with eight). The shell varies greatly from high- and low- spired helical coiled (snail-like) shells, planispirals (coiled in one plane) found in some Paleozoic marine and modern freshwater types, to cap-like limpets. There are many kinds of land slugs and marine sea-slugs which have lost their shell altogether, or have a small internal or vestigial shells beneath the mantle.
The head is well developed (only cephalopods have a more strongly developed head) with a mouth, tentacles, eyes, and other sense organs. In some advanced marine gastropods there is a tube-like structure - a proboscis - with which the animal can capture its prey. The head is more or less continuous with an elongate body. The digestive, reproductive, and other organs are coiled in a spiral which is enclosed in, and is protected by, the shell.
There is usually a muscular foot running the entire length of the ventral (lower) surface of the body. The animal creeps along the flat sole-like lower surface by small waves of muscular contraction, lubricated by slime from mucous glands.
The entire body (or head-foot) may be withdrawn into the last (largest or body) whorl of the shell by retractor muscles and in many marine and some land snails the opening may be closed with a horny or calcareous plug or door called an operculum, which is attached to the posterior dorsal surface of the body. This blocks the opening and protects the animal from predators or other dangers.
This ability to withdraw inside their shells and thus escape predators gave gastropods a great advantage, and they have been incredibly successful since their appearance in the Latest Cambrian right through to the present day.
The Gastropod Shell
Geometrically, the typical gastropod shell is a cone coiled round a central axis as a spiral. The reason the shell is coiled is simple - a high uncoiled shell would be impossible to carry because of its high center of gravity. therefore the shell is coiled and usually tilted. Most shells coil to right (dextral), although a few coil to left (sinistral), and there are even Palaeozoic types (some Euomphalida, especially macluritids) where the spire was "upside down" (hyperstrophic)
The following diagram shows the various parts of the gastropod shell.

The central axis of coiling is called the columella and at the anterior end of the shell forms the inner lip of the mouth opening, called the aperture. In neogastropods it may be drawn out to form with the anterior edge of the mouth a canal (anterior canal) sheathing the proboscis. There is also in some species, a notch between the outer lip and the body-whorl, this is called the posterior canal, for the inhalant siphon. The shape of the spiral varies considerably, as does the form of the aperture. The outer lip may be thickened or sharp and may or may not be indented with "teeth". At the apex of the spiral is the embryonic shell called the protoconch; it often has a different form and ornament from the adult shell [Macpherson and Gabriel 1962 p.26]
The single unique defining characteristic (or synapomorphy) of the gastropods is a process known as torsion. Even though not all gastropods are torted, those that are not have evolved from torted ancestors.
During the development of the planktonic larva, the visceral mass is horizontally twisted nearly 180° in a counterclockwise direction in relation to the lower part of the body. As a result, the mantle cavity, gills, anus, and two nephridopores are rotated from back to front, and are now located in the anterior position, above the head, and the body was no longer symmetrical.
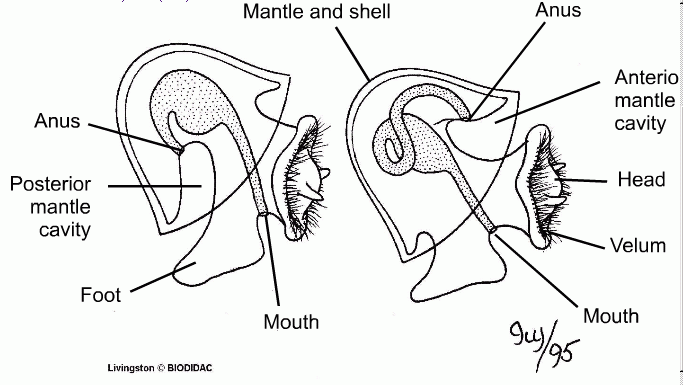
It is not quite clear why this first happened, more than five hundred million years ago. Walter Garstang in the 1920s proposed that it helps larval veliger pull its head inside its shell (and even wrote this as a poem: "Ballad of the Veliger or how the Gastropod got its Twist"!). Other explanations are that it helps the adult carry weight of shell, or it was an adult adaptation to protect the head and utilize the anterior water current.
Torsion obviously has benefits (including increased water current and allowing the animal to withdraw more deeply into the shell then would otherwise be possible), but it created a number of problems as well, the most serious of which was fouling. With the anus and nephridia in the front of the body, the animal would be dumping its waste on its head. This problem was solved by slit or cleft in the front of the shell, above the mantle cavity. pre-torted protogastropods already had a slit in their shell. This faced backwards in the pretorted mollusk, so when the entire shell was rotated through 180° it was at the front, and there was a ready-made solution (a perfect example of pre-adaptation). Thus the inhalant current continued to come in over the head and gills, but instead of making a U-turn and passing out the same way, it flowed up and out through the slit in the shell, taking the waste with it. [Barnes 1980, from Rollins and Batten 1968]
Note that torsion does not imply coiling. The first gastropods evolved from untorted ancestors like Crytolites that still had a coiled shell.
Gastropods have been a stable and constant component of the marine benthic ecosystems since the early Paleozoic. Unambiguous Gastropods first appear in the second half of the Furongian (Dolgellian Age), probably evolving from bellerophontiform "Tergomya" "monoplacophora") like Cyrtolites. Earlier spiral shells attributed to the class, and dating to the early and middle Cambrian, are now considered to be non-molluscan (e.g. Aldanella) or non-gastropod (either Helcionellid, tergomyan, or Paragastropod) mollusk.
Like the Cephalopods that also appeared at this time, Gastropods radiated very quickly, but unlike the Cephalopods made it through the end Cambrian mass extinction event with few casualties. These very early (latest Cambrian and earliest Ordovician) forms can in a variety of shell types, including bellerophontiform, discoidal (planispiral), low-spired anistrophical (conventional helically coiled spiral) and hyperstrophic ("upside down" spiral - Macluritina) coiled forms, loosely coiled, and high spired types.
Working out the evolutionary relationships of creatures long-gone is not easy. All these very early forms used to be included under the now redundant taxon "Archaeogastropoda". Also, Dr Wagner's cladistic analysis [Wagner 1999] has shown that many forms previously considered Vetigastropod (Pleurotomarid) are actually members of either the Euomphalida (probably Eogastropoda) or the Murchisoniida (probably Orthogastropoda) [Wagner no date]. The bellerophontiform gastropods he considers to be bi-phyletic, with one group perhaps ancestral to, and another an off-shoot of, a very early paraphyletic assemblage called the Sinuopeidae (latest Cambrian to middle of the Early Ordovician). Of course, it is also possible that the Sinuopeids evolved from torted bellerophontiforms, which in turn developed from untorted bellerophontiforms.
By Carboniferous times gastropod faunas were diverse. Not only were there a large number of marine forms, but some lineages had adapted to fresh water, and the first terrestrial forms appeared. These however were unrelated to the pulmonates of the Mesozoic to recent eras.
Like most other organisms, the gastropods were affected by end-Permian mass extinction event, and a number of groups died out at this time. Many more however survived through to the Mesozoic. A number of new groups flourished at this time, including the Nerineidae - a family of high-spired Mesozoic mesogastropods - and - later in the Mesozoic - the various Neogastropod taxa. The Jurassic and Cretaceous also saw the appearance and radiation of the Pulmonates or fresh-water and terrestrial forms. But the real acme of gastropod evolution was reached in the Cenozoic, continuing until the present, with the great success of the long-siphoned neogastropods which dominate today's gastropod fauna. [Clarkson, 1979, pp.163-64]
Gastropod systematics and phylogenetic understanding has undergone radical revision in the last few decades, and the old three-fold classification of Prosobranchia, Opisthobranchia, and Pulmonata, with the Prosobranchia divided into Archaeogastropoda, Mesogastropoda, and Neogastropoda, is no longer accepted by recent workers in this field, who have adopted a cladistic perspective based on more thorough recent research. The terms are however still found in older (and even fairly recent) books, and may still be used as generic categories - especially Linnaean categories. While there have been attempts to reconcile the current paradigm with the Linnaean hierarchy, such an approach is bound to fail, because these are two completely different methodologies. The following presents a cladistic summary:
Suggested revised Linnaean classification:
The following is suggested as a very tentative and provisional alternative to the traditional classification. In keeping with the convention of the Treatise authors of the volumes on Gastropoda and Cephalopoda, the suffix -ina is used for suborders, and -ida for orders.
Incertae Sedis (primitive forms - Prosobranchia / Archaeogastropoda in part)
Order "
Tropidodiscida"
("Bellerophontina" in part)
Order
Bellerophontida ("Bellerophontina" in part)
Order Cocculinida
(polyphyletic?, may be either Eogastropod or Orthogastropod) Subclass
Eogastropoda (primitive forms - Prosobranchia / Archaeogastropoda in part)
Order
Euomphalida
Order "Platycerida"
(normally a superfamily, see note)
Order
Patellogastropoda (Docoglossa) Subclass
Orthogastropoda (all other gastropods)
Infraclass unnamed
(Prosobranchia in part))
Superorder unnamed
(Archaeogastropoda in part)
Order Murchisoniida
Order Neritopsina
Order Neomphalida
Order Vetigastropoda
Superorder
Caenogastropoda
Order Architaenoglossa
Order Neotaenioglossa
Order Neogastropoda
Infraclass
Allogastropoda
Order Heterostrophia
Infraclass
Opisthobranchia
Order Architectibranchia
Order Cephalaspidea
Order Anaspidea
Order Thecosomata
Order Gymnosomata
Order Sacoglossa
Order Umbraculomorpha
Infraclass
Pulmonata
Order
Systellommatophora
Order
Basommatophora
"Order"
Actophila (= Archaeopulmonata?)
Order
Stylommatophora
note: the Platyceroidea are considered Trochid "Archaeogastropoda" (= Vetigastropoda) by Knight, et al., 1960 and the "Euomphalina" by Jeffery 2001 [following Wagner 1999] However they would seem to differ from ordinary Euomphalids as much as Patellogastropods do, so I have placed them in their own Linnaean) Order.
page uploaded 25 February 2003,
last modified ATW060227
checked ATW050720
original material by M. Alan Kazlev all other material © original authors or sources