Aculifera
Taxa on This Page
- Aculifera
- Phthipodochiton
- Mattheviidae
- Septemochiton
The Aculifera hypothesis
 Strict consensus tree from parsimony analyses showing inferred relationships among Aculifera and related taxa based on 101 most-parsimonious trees generated when brachiopod out-group characters were coded according to the brachiopod-fold hypothesis (Cohen et al. 2003). Lines in bold indicate living taxa; brachiopod (out-group) taxa are in grey; additional fossil outgroup taxa are indicated by asterisks. 'Polyplacophoran' groups Neoloricata, Palaeoloricata and Multiplacophora (Multiplac.) are labelled; molluscan crown group is indicated by the surrounding box. - cladogram and caption from Sigwart & Sutton 2007 fig 3. Note, Turrilepas Lepidocoleus, and Plumulites (shown at the upper left) are now known to be armoured annelids. However their close similarity with Paleozoic aculifera shows how closely related the two phyla are at thuis level (Vinther et al 2008 , Vinther & Briggs 2009) Strict consensus tree from parsimony analyses showing inferred relationships among Aculifera and related taxa based on 101 most-parsimonious trees generated when brachiopod out-group characters were coded according to the brachiopod-fold hypothesis (Cohen et al. 2003). Lines in bold indicate living taxa; brachiopod (out-group) taxa are in grey; additional fossil outgroup taxa are indicated by asterisks. 'Polyplacophoran' groups Neoloricata, Palaeoloricata and Multiplacophora (Multiplac.) are labelled; molluscan crown group is indicated by the surrounding box. - cladogram and caption from Sigwart & Sutton 2007 fig 3. Note, Turrilepas Lepidocoleus, and Plumulites (shown at the upper left) are now known to be armoured annelids. However their close similarity with Paleozoic aculifera shows how closely related the two phyla are at thuis level (Vinther et al 2008 , Vinther & Briggs 2009) |
The name Aculifera is given to a clade of molluscs recovered through molecular phylogeny and some paleontological analyses (Sigwart & Sutton 2007 Kocot et al 2011, Smith et al 2011, Vinther et al 2011). It includes all molluscs that either primitively lack a shell (as opposed to having secondarily lost their shell) (aplacophora) or have a series of plates instead of a single shell (polyplacophora). These were originally called Amphineura, a class that was rejected be cause there are no morphological features that unite them to the exclusion of other molluscs. The group was restored, but under a different name, following some morphological studies, but was never popular until being recovered by recent molecular and cladistic analyses. As currently defined, the Aculifera includes the eight-plated Polyplacophora (chitons) as well as the shellless and either primitive or degenerate Caudofoveata (=Chaetodermomorpha) and Solenogastres (=Neomeniomorpha), along with their fossil relatives. In terms of taxonomic rank, Aculifera is one of the two subphyla of the molluscs, the other being the Conchifera (shell-bearers), which includes the majority of molluscs. The cladogram on the right provides one possible phylogeny, and assumes a monophyletic aculifera and mollusca, which emerges from a paraphyletic halwaxiida and brachiopoda.
A similar phylogeny, shown below, is provided by the sketch of early molluscan evolution by Dr Jakob Vinther and co-workers. In this cladistic diagram, once again, aplacophorans emerge through the secondary loss of shell (and perhaps also the foot) from a paraphyletic assemblage of "paleoloricates". Illustrated are two early Paleozoic genera, the bizarre worm-like Acaenoplax, which seems to possess a combination of aplacophoran and chiton attribites, and the very basal spiny plated Cambrian form Matthevia
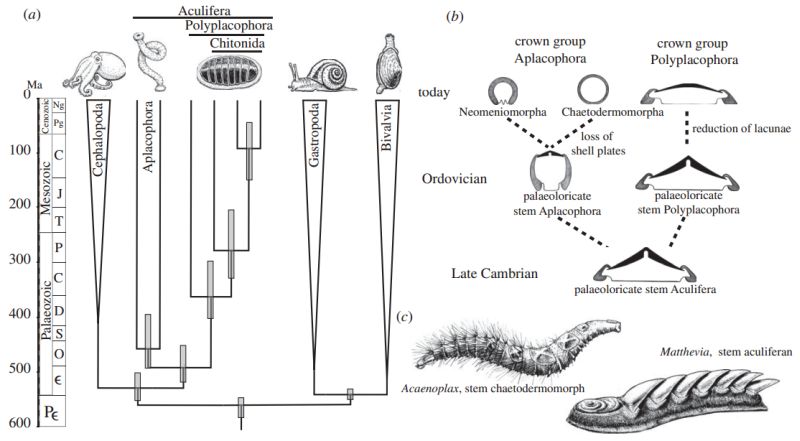 Aculiferan and molluscan evolution. (a) Timetree of aculiferans assessed with Bayesian relaxed molecular divergence estimation). Grey bars are 95% credibility intervals. (b) Morphological transitions in aculiferan evolution in relation to the foot, shell and mantle, based on molecular evidence presented herein and partly on previous morphological cladistic hypotheses [19,22]. The ancestral aculiferan was a form with a broad ciliary foot and a shell with lacunae (Paleoloricate). The molecular clock analysis and the fossil record indicate that in the Ordovician, the Aculifera had diverged into a stem group aplacophoran with a reduced foot and a laterally compressed body; fossil evidence demonstrates that this form was of palaeoloricate morphology with lacunae in the shell plates. Stem group polyplacophorans also retained the plesiomorphic palaeoloricate condition at this time. The aplacophorans diverged early into the chaetodermomorphs and the neomeniomorphs. Chitons reduced the lacunae in the shells and developed a more dorsoventrally flattened aspect and a smoother dorsal surface. (c) Reconstructions of two important fossil forms: Acaenoplax hayae (an aplacophoran relative) and Matthevia (the earliest known stem group aculiferan with multiple overlapping plates). -Diagram and caption from Vinther et al 2011 fig 4
Aculiferan and molluscan evolution. (a) Timetree of aculiferans assessed with Bayesian relaxed molecular divergence estimation). Grey bars are 95% credibility intervals. (b) Morphological transitions in aculiferan evolution in relation to the foot, shell and mantle, based on molecular evidence presented herein and partly on previous morphological cladistic hypotheses [19,22]. The ancestral aculiferan was a form with a broad ciliary foot and a shell with lacunae (Paleoloricate). The molecular clock analysis and the fossil record indicate that in the Ordovician, the Aculifera had diverged into a stem group aplacophoran with a reduced foot and a laterally compressed body; fossil evidence demonstrates that this form was of palaeoloricate morphology with lacunae in the shell plates. Stem group polyplacophorans also retained the plesiomorphic palaeoloricate condition at this time. The aplacophorans diverged early into the chaetodermomorphs and the neomeniomorphs. Chitons reduced the lacunae in the shells and developed a more dorsoventrally flattened aspect and a smoother dorsal surface. (c) Reconstructions of two important fossil forms: Acaenoplax hayae (an aplacophoran relative) and Matthevia (the earliest known stem group aculiferan with multiple overlapping plates). -Diagram and caption from Vinther et al 2011 fig 4 |
Unlike the mammalian molecular clock, with its non-existent Mid-Late Cretaceous and Mid Tertiary placental radiations , this seems to be an instance where the molecular clock supporerts stratigraphy and vice-versa. In keeping with the phylogeny in Vinther et al 2011, the above cladogram presents aculiferans and cephalopods as sister taxa. As there is no morphological or paleontological support for this molecular-only hypothesis, we have chosen instead the phylogeny of Kocot et al 2011, according to which cephalopods evolved from monoplacophora (and the two are hence sister taxa, as in the molecular phylogeny of Smith et al 2011
Fossil Record
If Early to Middle Cambrian Sachitids are aculiferan ((Vinther 2009).) then the group appears at the start of the Paleozoic, which makes sense given the similar antiquity of the conchifera (higher molluscs). It may even be that halkeriids are aculiferans (Vinther & Nielsen 2005). The implication is that there is an unbroken continuum from basal halwaxiids to molluscs proper, and that some halwaxiids are basla molluscs. and even basal aculifrean; although if teh aculifera are paraphyletic, as we believe, than that last phrase is redundant) In any case, these early molluscs and proto-mo;;uscs seme to have bene common and widespread during the early Cambrian.. By the Late Cambrian they had goven rise to an accentric assemblage of Mattheviids which represent the predominant Late Cambrian and Ordovician aculiferan type.
During the Silurian there was a radiation of heloplacids, which combined the characteristics of chitons and aplacophoran molluscs. Multiplated skeletons in paleoloricates and heloplacid aplacophorans represent either parallel evolution (Cherns 2007) or the plesiomorphic state of dorsal armour (Sigwart & Sutton 2007, Vinther et al 2011), or both, and is not overlapping, unlike true chitons. Peaks of aculiferan evolutionary diversity in early Ordovician, mid-Silurian and early Carboniferous times correspond to periods with development of low latitude carbonate shelves. True chitons (Neoloricates) appear in the Devonian , and have an additional shell layer that adds articulatory plates. However the fossil record of the group at this time remains poorly known (Cherns 2007)
Are the Aculifera paraphyletic?
A challenge with molluscan phylogeny, as with phylogeny in general, is finding an overall explanation that can accomodate the more often contrary and less often mutually supporting hypothesis from morphological, functional, developmental, paleontological, stratigraphic, cladistic, and molecular methodologies. Too often only one methodology is used (the tendency now is to rely mostly or solely on molecular data) and contrary hypotheses using other methodologies is rejected. For example Salvini-Plawen 1980 and elsewhere, and Haszprunar 2000 argue on morphological grounds that the aplacophoran groups and the polyplacophora represent a gradation of primitive types leading to the Conchifera or higher molluscs. But in the 2000s through to 10s , cladistic and molecular studies (Sigwart & Sutton 2007, Kocot et al 2011, Smith et al 2011, Vinther et al 2011) reject a paraphyletic aplacophora in favour of the Aculifera hypothesis. It is asusmed that the more recent papers are more correct, and that the earlier work is outdated. But this is a misleading premise, because very different methodologies are used in each case, each with different phylogenetic signals, and it is not possible to refute one methodology using a totally unrelated methodology (e.g. morphology-based cladistics cannot be disproved by molecular sequencing, and vice-versa). There is also no way of knowing when a new paradigm will emerge, or new evidence uncovered, causing a toal turn around. For example in the 1950s 60s and 70s, text books regularily referred to the Amphineura (=aplacophora + polyplacophora). Following the work of Salvini-Plawen and others, the Amphineura was rejected and replaced by two or three unrelated classes. Now the tendency is for the aplacophora and polyplacophora to be again grouped together in a single group, now called Aculifera, despite there being very little in morphological terms that these various groups have in common, other than plesiomorphic molluscan or spiralian characteristics, such as spicules and multiple dorsal shell plates, both features shared with halwaxiids but lost in conchifera.
Although both molecular and paleontological-statistical cladistic data (Sigwart & Sutton 2007, Vinther et al 2011) support the Aculifera hypothesis (see cladograms above), the problem with a monophyletic aculifera is that it leaves the ancestry of the Conchifera hanging in the air. Since we do not wish to multiply entities unnessarily (William of Occam), iit seems much more parsimonious to reject the ghost lineages and simply assume that the Aculifera - which are morphologically intermediate between halwaxiids and conchifera - are a paraphyletic group. Their apparently monophyletic status can be explained by the loss or transformation of the original aculiferan synapomorphies in their conchiferan descendents (the same applies to Cambrian arthropods (Walossek & Müller 1998 pp.186f). If the same happened in the genome as well as the gross morphology, molecular phylogeny would likelwise give a "false monophyly" result (as with the cyclostome jawless fish, which are morphological a paraphyletic or polyphyletic gradation (as conclusively shown by Janvier) but in molecular phylogeny uniformly appear as monophyletic).
On the other hand, if the aculifera predate and give rise to the conchifera, it is necessary to explain why the earliest known fossil aculiferans are no older than the Late Cambrian, whereas the Conchifera were already diverse and numerous by the Early Cambrian. So hybrid forms such as Matthevia, which seem to combine attributes of chitons and monoplacophora, rather than being ancestral to the monoplacophora, may have evolved from them. Or the features the two groups have in common may simply be shared ancestral traits. The lineage represented by the Silurian spiny worm-like Acaenoplax may also belong here, or it may be closer to crown group aplacophora. It is also quite possible that Halkieria-like animals were stem mollusca (or even aculiferan (Vinther & Nielsen 2005, Vinther 2009), although that would make the latter group paraphylertic), and that they were the ancestors both of Mattheviid type multi-placophorans and conchiferan monoplacophora, both of which can be considered transitional forms. MAK120613
Descriptions
Aculifera Hatscheck, 1891
Phylogeny: paraphyletic Halwaxiida/ basal Mollusca : Conchifera + * : paraphyletic Mattheviidae + Phthipodochiton + Septemochiton + (Heloplacidae + (Caudofoveata + Solenogastres)) + Polyplacophora
spicules, multiple dorsal paltes or spines, larval or adult metamerism
Comments: Two possible classifications of Aculifera as a subphylum: Aculifera includes the Aplacophora (Caudofoveata and Solenogastres) and Polyplacophora (chitons), with all other molluscs being included in subphylum Conchifera (shell-bearers) (Hatscheck in Blumrich, 1891; Scheltema, 1993) or it including only the Aplacophora, with the polyplacophora going in the subphylum Placophora, and the Conchifera as previously described (Stasek, 1972). Haszprunar 2000, and Glaubrecht, Maitas & Salvini-Plawen, 2005) all would support a phylogenetic relationship like Solenogastres + (Caudofoveata + (Polyplacophora + Conchifera)) making the Aplacophora (and therefore Aculifera) paraphyletic. (Gofas (nd). In contrast, Sigwart & Sutton 2007 Kocot et al 2011, Smith et al 2011, Vinther et al 2011 all support a monophyletic Aculifera. Because groupings of molluscan classes into clades is highly controversial (for a review of competing hypothesis see Steiner & Dreyer (2003) , the various molluscan classes are better treated as an unresolved polytomy (Gofas (nd)) or the Conchifera and the three aculiferan classes can be considered an unresolved quadotomy. The Solenogastres and Caudofoveata can be derived from Paleozoic Mattheviids or Phthipodochiton or similar (Sigwart & Sutton 2007). Supporting the validity of the aculifera is the discovery that extant Solenogastre and Caudofoveatan embryology exhibits a chiton-like 7 fold iteration on the dorsal surface in late stages of trochophore larvae, which is lost during metamorphosis (Vinther ref). It is also possible that the multiplate chiton condition may be a primitive molluscan characteristic (related to annelid segmentation) which was lost with monoplacophorans (basal conchifera). If aplacophora and polyplacophora diverged in the Early Ordovician (Vinther et al 2011) than the Aculiferaare a monophyletic clade, although taxa such as Hemithecella and Matthevia with monoplacophora-like features could still be late surviving stem groups whose ancestors must have evolved during the very earliest Cambrian and would no doubt be among some of the problematic small shelly forms or Halkeriid-like animals. MAK120608
Mattheviidae Walcott, 1885 : Calceochiton, Chelodes, Eukteanochiton, Hemithecella, Matthevia, Robustum, Spicuchelodes (ref: Paleobiology Database)
Higher taxa and partial synonyms: Matthevina Fisher 1962; Chelodina Runnegar et al. 1979; Paleoloricata Cherns 1998.
Late Cambrian to Early Devonian
Phylogeny: Aculifera : Phthipodochiton + Septemochiton + (Heloplacidae + (Caudofoveata + Solenogastres)) + Polyplacophora + *
Characters: massive, elongate, non-overlapping valves, monoplacophoran-like shells and muscle scars. Paraphyletic assemblage
Comments: Either basal aculiferan, intermediate between aculifera and monoplacophora, or both.
Matthevia is a Cambrian mollusc, with repeated monoplacophoran-like shells projecting dorsally as vertical plates or spines. Originally it was proposed that chitons arose when these tall shells began to overlap (Runnegar & Pojeta 1974). Now that aculiferans rather than monoplacophorans seem to be the most primitive molluscs, it may be that instead these plates merged, or one developed at the expense of the others. There are distinct head, 'centre', and tail valves, which occur approximately in the ratio 1:5:1 — suggesting a seven-plated configuration Wikipedia, MAK120608
Hemithecella is a problematic multiplated late Cambrian and Early Ordovician genus, with muscle scars identical to a monoplacophorans, unlike the musculature of the chitons. It therefore cannot be included in the latter group (Stinchcomb & Darrough 1995, Pojeta et al 2010). Wikipedia
Vendrasco and Runnegar argue that mattheviids are polyplacophoran on the basis of shared valve characters such as including granules, an apical shelf, a thin anterior margin, bilateral symmetry, three valve types, and a shell layer perforated with canals. The mattheviids themselves constitute a gradation between classic forms such as Matthevia variabilis with non-overlapping, spiky valves, and more traditional paleozoic chitons such as Chelodes (Vendrasco & Runnegar 2004). MAK120607
SeptemochitonBergenhayn, 1955
Phylogeny: Basal Aculifera
Comments: Originally sole species of monotypal Family Septemochitonidae and Suborder Septemchitonina. Seven rather than the usual eight valves; phylogenetic position uncertain; the cladogram by Sigwart & Sutton 2007 makes Septemochiton the sister taxon of the Mattheviid Robustum. Hoare & Pojeta 2006 define the Septemochitonida Bergenhayn, 1955 as "Paleoloricates with intermediate plates differentiated into central and lateral areas." and include a number of families and genera. They fgive the range as Ordovician to Devonian MAK120607
Phthipodochiton thraivensis (Reed, 1911)
Synonym: Helminthochiton thraivensis Reed, 1911
Horizon: Lady Burn Starfish Beds, Ordovician of Girvan, southwest Scotland
Phylogeny: Basal Aculifera
Comments: Worm-like body plan, lacks a true foot despite bearing polyplacophoran-like valves: a spicular girdle wraps entirely around the ventral surface of the animal, implying that a polyplacophoran like foot was absent, although traces of a narrow solenogastre-like median pedal groove is possiblly evident in the fossils (Sigwart & Sutton 2007, Sutton & Sigwart 2012). That means that these animals either had not yet acquired the creeping molluscan foot or had aleady secondarily lost it. The valves are typically palaeoloricate in aspect, but differ in detail from all other palaeoloricate genera, hence the new generic name Sutton & Sigwart 2012). Carnivorous, fed on crinoids (Donovan et al 2011); has an apparent mosaic of aplacophoran and polyplacophoran features, and shows that at least some other palaeoloricates possessed a worm-like "armoured aplacophoran" body plan, in contrast to the limpet-like body plan of extant Polyplacophora Sutton & Sigwart 2012); the latter may simply be an adapatation to a specialised tidal-zone ecological niche among this ancient group MAK120612.