| Palaeos: | Bones: The Braincase | |
| Vertebrates | The Sphenethmoid |
| Page Back | Unit Back | Unit Home | Unit References | Unit Dendrograms ("Cladograms") |
Glossary | Taxon Index |
| Page Next | Unit Next | Vertebrates Home | Vertebrate References | Vertebrate Dendrograms | Bones | Time |
|
Bones
|
Braincase
|
Sphenoid Region
Overview |
 Anatomy: The sphenethmoid, sometimes called the ethmosphenoid, is something between a
region and a bone. In primitive gnathostomes, the embryonic trabeculae (see Overview) fuse to form a single ossification which includes the
entire braincase from the ethmoid plate, anteriorly, to the otic region
posteriorly. This region is marked by the orbits, laterally, and often the
articulation between the braincase and the palatoquadrate. Its posterior extent
was coincident with the original anterior limit of the notochord and the ventral
fissure. Ventrally, the ethmosphenoid normally contains foramina for the
internal carotid artery and the Vth cranial (trigeminal) nerve, or at least the
profundus branch of that nerve. The sphenethmoid also usually bore the
basicranial articulation, the old major articulation between the palate and the
braincase.
Anatomy: The sphenethmoid, sometimes called the ethmosphenoid, is something between a
region and a bone. In primitive gnathostomes, the embryonic trabeculae (see Overview) fuse to form a single ossification which includes the
entire braincase from the ethmoid plate, anteriorly, to the otic region
posteriorly. This region is marked by the orbits, laterally, and often the
articulation between the braincase and the palatoquadrate. Its posterior extent
was coincident with the original anterior limit of the notochord and the ventral
fissure. Ventrally, the ethmosphenoid normally contains foramina for the
internal carotid artery and the Vth cranial (trigeminal) nerve, or at least the
profundus branch of that nerve. The sphenethmoid also usually bore the
basicranial articulation, the old major articulation between the palate and the
braincase.
However, in early tetrapods, this portion of the braincase began to take on a rather different character. With the development of skulls constructed of strong dermal bones, the importance of the sphenoid was reduced. The basicranial articulation was no longer the anchor for the palate, and the palatoquadrate was functionally replaced by its dermal anlagen, the maxillae. By the level of the amniotes, the sphenethmoid had disappeared altogether. Instead, the midbrain was roofed by dermal bones, and the principle sphenoid ossifications became a series of bar-like bones which supported the brain ventrally and sometimes laterally. These are the various sphenoid bones which will be dealt with separately. The remainder of the anterior sphenoid region was unossified.
 The bottom line of all this is that the Standard Condition for the sphenethmoid
cannot be defined with reference to our usual informant, Bob the Basal Amniote.
Bob simply doesn't have a sphenethmoid. Instead, it is convenient to define the
Standard Condition by reference to some, vaguely dipnomorphic,
sarcopterygian whom we will call
"Judy," after another primitive, schizocephalic blockhead not
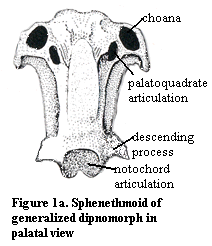
otherwise discussed in these Notes. Judy's isolated sphenethmoid is shown in
palatal view in Figure 1a. (Figures 1a, 1b and 1c are modified
from illustrations of Gogonasus andrewsae in Long,
1995). As in many sarcopterygians, Judy's sphenethmoid is confluent with the
ethmoid and nasal capsules. This the choanae lead dorsally right into the nasal
capsules. The orbit and jaw adductors fit into the 'C'-shaped cavities on either
side of the long sphenoid stem region.
The bottom line of all this is that the Standard Condition for the sphenethmoid
cannot be defined with reference to our usual informant, Bob the Basal Amniote.
Bob simply doesn't have a sphenethmoid. Instead, it is convenient to define the
Standard Condition by reference to some, vaguely dipnomorphic,
sarcopterygian whom we will call
"Judy," after another primitive, schizocephalic blockhead not
otherwise discussed in these Notes. Judy's isolated sphenethmoid is shown in
palatal view in Figure 1a. (Figures 1a, 1b and 1c are modified
from illustrations of Gogonasus andrewsae in Long,
1995). As in many sarcopterygians, Judy's sphenethmoid is confluent with the
ethmoid and nasal capsules. This the choanae lead dorsally right into the nasal
capsules. The orbit and jaw adductors fit into the 'C'-shaped cavities on either
side of the long sphenoid stem region.
In Figure 1b, we add ghost-like images of Judy's premaxilla, parasphenoid and the otico-occipital region of the braincase. The premaxillae fit tightly onto the anterior end of the ethmoid region and add a tooth row. As discussed at The Premaxilla, the premaxilla continues its contact with the nasal capsules and becomes involved with the external nares as well. The parasphenoid is a tooth- or denticle-covered plate which is attached to the sphenoid stem. The otico-occipital braincase attaches to the posterior. Note how the descending processes of the sphenethmoid hook onto anterolateral facets of Judy's prootic to form a kind of hinge joint on which the intercranial joint can flex. The otico-occipital portion is not only more massive than the sphenethmoid, but it is stabilized by the notochord which enters through the posterior "stem" region, runs all the way through the otico-occipital braincase and terminates at a socket joint at the end of the sphenethmoid, providing yet another mobile contact which allows the sphenethmoid to flex downward. The muscular workings of the intracranial joint are discussed in more detail (with yet another complicated diagram in lateral view) in connection with the main entry on porolepiforms.
 Finally,
we are ready to add some additional palatal roofing bones. These are shown in Figure
1c. The maxilla extends the skull outward from the sphenethmoid to make
room for Judy's big mouth and, perhaps more importantly, a larger, more
posterior adductor chamber for the all-important jaw-closing muscles. Her
palatines supply an additional row of teeth and the pterygoids almost complete
the roof of the palate. This is as far as we will take the assembly, but the
palate is still incomplete. For example, the blank area between the choanae
would be covered by the vomers. The second tooth row might be extended by
toothed ectopterygoids. A second layer of denticle-bearing dermal bones, such as
entopterygoids and parasphenotic plates, might cover all or part of the
pterygoids.
Finally,
we are ready to add some additional palatal roofing bones. These are shown in Figure
1c. The maxilla extends the skull outward from the sphenethmoid to make
room for Judy's big mouth and, perhaps more importantly, a larger, more
posterior adductor chamber for the all-important jaw-closing muscles. Her
palatines supply an additional row of teeth and the pterygoids almost complete
the roof of the palate. This is as far as we will take the assembly, but the
palate is still incomplete. For example, the blank area between the choanae
would be covered by the vomers. The second tooth row might be extended by
toothed ectopterygoids. A second layer of denticle-bearing dermal bones, such as
entopterygoids and parasphenotic plates, might cover all or part of the
pterygoids.
Dorsally, the sphenethmoid is in contact with the dermal bones of the facial, orbital and vault series. However, in dipnomorphs, these bones tend to be rather non-standard (see figure at Dipnoi), so that the articulations are not what one might normally expect.
Functions: In the Standard Condition, the sphenethmoid is associated with the following, fairly obvious, functions:
1) Formation of the anterior braincase. The sphenethmoid is intimately connected with the olfactory nerves, the entire optic apparatus and the fore- and mid-brain.
2) Anterior articulation of the palatoquadrate. Technically, this is part of the ethmoid, but the two are confluent in the Standard Condition.
3) Medial support of the palate. In sarcopterygians, including basal tetrapods, the mouth is very large. It is stable only because the sphenethmoid (and its later successors) provide a roof beam which supports the center of the palate.
4) Intracranial articulation. The sphenethmoid is the moving part of the intracranial joint. In many sarcopterygians, the joint gradually became immovable, or only slightly flexible as in the living actinistian, Latimeria. However, it seems to have played an important role in the evolution of the structure of the sarcopterygian head which the tetrapods inherited.
 Phylogeny:
Phylogeny:
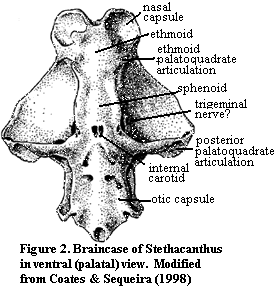
1) Basal gnathostomes: There will always be a sphenoid, if for no other reason than that there must be something between the nose and the otic capsules. Figure 2 shows as primitive a gnathostome braincase as we are likely to know for some time. This is the early symoriid chondrychthian, Stethacanthus. In spite of the vast phylogenetic gap between Stethacanthus and dipnomorphs, many of the sphenethmoid landmarks are recognizable. These include the palatoquadrate articulation at the posterior boundary of the ethmoid, the long, thin sphenoid region, and the trigeminal and internal carotid foramina. In fact, not a great deal separates the sphenethmoid areas of Stethacanthus and the Standard Condition except for the absence of both endochondral bone and the intracranial fissure in the former. Not surprisingly, the sphenethmoid of the basal sarcopterygian Psarolepis is more or less intermediate between the two. Yu (1998).
2) Basal tetrapods: The advanced sarcopterygian Eusthenepteron also has the Standard Condition, except that the trigeminal nerve foramina are all posterior to the intracranial division. Since the Vth nerve elements would then have to cross the intracranial divide, this suggests (as do some subtle changes in geometry) that the intracranial joint was less flexible. The sphenethmoid region as a whole already shows a trend toward closer integration with the dermal bones dorsally, with a more open structure on the dorsal surface of the braincase. By contrast, the ventral surface of the sphenethmoid is much broader in proportion to the total width of the head, looking more like a beam than a hammer.
 With
the move onto land, tetrapods* lost the constant support of an aqueous medium. As
a result, there was considerable adaptive advantage to reducing the mass (and so
the moment arm) of anterior element of the head. The labyrinthodonts still had
fairly elaborate dermal shoulder girdles which could allow the arms to support
the back of the head directly. However, the anterior elements were unsupported
and enormous energy would be required to hold them off the ground -- similar to
the difference between holding a bowling ball at arms' length, rather than close
to the body. As a result, it is not surprising that temnospondyls
and other stem tetrapods rearranged their skulls significantly. Thus the
basicranial articulation was moved to the otico-occipital region, and the
ethmosphenoid became a largely unossified element located half-way out on a
long, thin beam created by the cultriform process of the parasphenoid which
passed through the middle of two large, empty (of bone) "interpterygoid
vacuities" The general scheme can be seen in the figure of Laidleria,
a moderately derived temnospondyl, at right. Warren
(1998). Here, the otico-occipital unit would be supported by the posterior
body of the parasphenoid, while the remaining sphenethmoid would be a small
element attached to the long anterior cultriform process of the parasphenoid.
With
the move onto land, tetrapods* lost the constant support of an aqueous medium. As
a result, there was considerable adaptive advantage to reducing the mass (and so
the moment arm) of anterior element of the head. The labyrinthodonts still had
fairly elaborate dermal shoulder girdles which could allow the arms to support
the back of the head directly. However, the anterior elements were unsupported
and enormous energy would be required to hold them off the ground -- similar to
the difference between holding a bowling ball at arms' length, rather than close
to the body. As a result, it is not surprising that temnospondyls
and other stem tetrapods rearranged their skulls significantly. Thus the
basicranial articulation was moved to the otico-occipital region, and the
ethmosphenoid became a largely unossified element located half-way out on a
long, thin beam created by the cultriform process of the parasphenoid which
passed through the middle of two large, empty (of bone) "interpterygoid
vacuities" The general scheme can be seen in the figure of Laidleria,
a moderately derived temnospondyl, at right. Warren
(1998). Here, the otico-occipital unit would be supported by the posterior
body of the parasphenoid, while the remaining sphenethmoid would be a small
element attached to the long anterior cultriform process of the parasphenoid.
3) Lepospondyls and Lissamphibia: Microsaurs adopted more or less the same design as temnospondyls, but the evolutionary constraints they faced were not as severe because of their relatively small size and aquatic habit. In the microsaur Rhynkconkos, for example, the sphenethmoid loses the basicranial articulation and the trigeminal nerve, as in temnospondyls. Likewise it is restricted to a narrow section dorsal to the cultriform process. However, it is still a long, fairly substantial element which is in direct contact with the basisphenoid and the otico-occipital section of the braincase.
 Caecilians,
generally speaking, took the microsaur body plan and consolidated it for their
difficult, fossorial mode of life. As shown in the figure, the sphenethmoid
makes a modest comeback as a major cranial bone, but its former territory is
split with a pleurosphenoid ossification. The caecilians have also developed a
unique paired infrafrontal process of the sphenethmoid which may help support
the dermal skull bones against the considerable forces generated by digging.
Caecilians,
generally speaking, took the microsaur body plan and consolidated it for their
difficult, fossorial mode of life. As shown in the figure, the sphenethmoid
makes a modest comeback as a major cranial bone, but its former territory is
split with a pleurosphenoid ossification. The caecilians have also developed a
unique paired infrafrontal process of the sphenethmoid which may help support
the dermal skull bones against the considerable forces generated by digging.
A rather similar arrangement occurs in frogs. However, the anuran ethmoid plate widens out considerably at the anterior of the skull, so that the whole external aspect of the skull is changed. The premaxilla and maxilla look like a large arch supported at the apex by a pillar made up of the cultriform process of the parasphenoid with its attached sphenethmoid (and a more posterior ossification which is likely a pleurosphenoid).
4) Amniotes: There are occasional references to ossified sphenethmoids in the amniote literature. See, e.g., Currie (1997) (dinosaurs). However I have not yet found anything specific. At least generally speaking, the sphenethmoid is not ossified in amniotes.
ATW 010504.
| Page Back | Unit Home | Glossary | Page Top | Page Next |
checked ATW060127