| Palaeos: | Bones: The Braincase | |
| Vertebrates | The Opisthotic |
| Page Back | Unit Back | Unit Home | Unit References | Unit Dendrograms ("Cladograms") |
Glossary | Taxon Index |
| Page Next | Unit Next | Vertebrates Home | Vertebrate References | Vertebrate Dendrograms | Bones | Time |
|
Bones
|
Braincase
Overview |
Otic Region
Demosphenotic: see Overview
(Dermosphenotic) |
The opisthotic is, as the name might suggest, behind the prootic. Conceptually, it is the posterior half of the otic capsule. In this capacity, it is a rather boring bone. Usually, it is hopelessly fused to other things, forming yet another set of inscrutable speed bumps and pot holes on the braincase and providing the basis for yet another set of hopelessly inappropriate anatomical terms in an extinct language. In this case the terms usually begin with crista, tecta, sella, or (who knows?) taiga, tundra, and the like. Braincase terminology is somewhat fractal. Each level of obscure anatomical referents turns out to be made up of parts and variants with even more eldritch anatomical names. Just as the essentially Greek braincase regions are made up of little Latin structures, there is probably an entire vocabulary of component substructures which is represented in Babylonian pictograms, the true meaning of which is revealed only to a secret hereditary caste of anatomical hierophants.
Fortunately, the opisthotic has another and more democratic nature. Like the premaxilla, it has been recruited to perform a variety of external functions whose nature is open and accessible to all. Almost every student who has looked a figure including the occiput -- the bones at the back of the head -- has seen the "paroccipital process" and has asked, plaintively, "the paroccipital process of what?" Here, all will be revealed. The paroccipital process is actually a process of the opisthotic. However, the process is a truly public structure and other elements tend to get involved, including the exoccipitals, supraoccipital, and even the prootic. In fact, there are "paroccipital processes" that do not even involve the opisthotic. Thus the structure tends to be referred to as if it were a separate bone.
Accordingly, the opisthotic has a two-fold personality. On the one hand, it is an obscure and secretive braincase element. On the other, a flamboyant and out-going external structural element of the occiput. We will concentrate primarily on the latter aspect. It is not only more accessible, but has recently been the subject of an odd and interesting study by Dr. David Berman of the Carnegie Museum, better known as the co-discoverer of the Bounding Bolosaur -- Eudabimus. Berman et al. (2000). But Berman (2000) is a very different kind of paper. Here, Berman carefully develops his theory of the evolution of the amniote occiput. Although it has all the appropriate cladistic tools and terminology, it is essentially a pre-cladistic work in the best tradition of Huxley's On the Theory of the Vertebrate Skull. That is, it begins with a few key developmental and anatomical insights and works them through numerous taxa to arrive at a unified picture of evolution at work. This kind of essentially intuitive work is still critical, and always will be. The cladistic analysis of numerical characters is the almost magical rocket engine driving contemporary paleontology. Still, someone has to point the thing in the right general direction and send us the postcards telling us where it has been. Berman (2000) is that kind of paper. Unfortunately, Berman's paper involves some aggressive reinterpretation of a number of well known taxa. So, it is a hard paper to evaluate. Nevertheless, it contains a good deal of information and raises a good many useful issues.
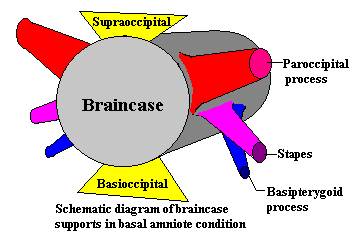 As
usual, Bob, the Basal Amniote, is the
standard bearer of the Standard Condition. However, even before we begin our
customary visit to Bob, it is useful to look at a schematic. This is a sort of
roadmap. If we model the braincase (very inaccurately) as a cylinder, we see
that it is braced primarily towards the posterior end. Ventrally, it is
supported by the basioccipital, which usually articulates with the pterygoids.
Dorsally, the supraoccipital braces the braincase against the skull table,
usually through some combination of the parietals, postparietals and tabulars.
From the opisthotic at the rear of the otic capsule, the paroccipital process
extends laterally, usually in combination with the exoccipitals but often
including other elements. The paroccipital process typically terminates at the
squamosal. Finally, the stapes creates a ventrolateral brace, joining the
braincase with the quadrate at the ventrolateral corner of the occiput.
As
usual, Bob, the Basal Amniote, is the
standard bearer of the Standard Condition. However, even before we begin our
customary visit to Bob, it is useful to look at a schematic. This is a sort of
roadmap. If we model the braincase (very inaccurately) as a cylinder, we see
that it is braced primarily towards the posterior end. Ventrally, it is
supported by the basioccipital, which usually articulates with the pterygoids.
Dorsally, the supraoccipital braces the braincase against the skull table,
usually through some combination of the parietals, postparietals and tabulars.
From the opisthotic at the rear of the otic capsule, the paroccipital process
extends laterally, usually in combination with the exoccipitals but often
including other elements. The paroccipital process typically terminates at the
squamosal. Finally, the stapes creates a ventrolateral brace, joining the
braincase with the quadrate at the ventrolateral corner of the occiput.
These are very broad generalizations, with numerous exceptions. However, they give us some basic landmarks. Landmarks are frequently necessary because, unlike more orderly bones, the opisthotic doesn't seem to grow so much as it runs. That is, it behaves like melted plastic and thoughtlessly fails to leave visible sutures. For that reason, three other historic landmarks merit a very brief side trip:
 (a)
The posttemporal fenestra: This structure is of greatest
significance in non-amniotes. It is an opening in the back of the skull between
the paroccipital process and the bones of the skull table. In amniotes, it tends
to get covered over or obliterated.
(a)
The posttemporal fenestra: This structure is of greatest
significance in non-amniotes. It is an opening in the back of the skull between
the paroccipital process and the bones of the skull table. In amniotes, it tends
to get covered over or obliterated.
(b) The jugular foramen: Another landmark of occasional interest is the jugular foramen -- a small hole where the jugular vein exits the braincase. It generally marks the exit of the Xth and XIth nerves, as well. This jugular foramen is normally located along the articulation between the opisthotic and the exoccipitals.
(c) The fenestra ovalis (a/k/a fenestra vestibulae): This is the sensory "port" for the inner ear, as described in more detail at The Ear. It is frequently found along the prootic-opisthotic boundary.
With this roadmap, we can start to get into Bob's head. [1] In the case of Bob, the paroccipital process incorporates the exoccipitals. As mentioned above, the paroccipital process frequently includes other elements such as the supraoccipital or prootic. Bob has a very large posttemporal fenestra. Frequently much of the fenestra is obscured because the margin of the tabular which forms the dorsal border of Bob's fenestra develops into an opisthotic process which contacts the dorsal margin of the paroccipital process. A final variation is found in a number of basal amniotes, including the original model for Bob, Paleothyris. In these forms, the paroccipital process never reaches the squamosal. Instead, the process is present as a large, plate-like slab of bone lying between the supraoccipital and skull table, on one side, and the ventral margin of the occiput, on the other.
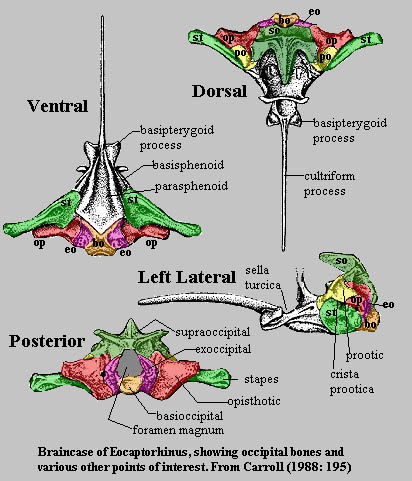 Before
leaving the standard condition, it is worthwhile to take a careful look at the
appearance of the braincase In another early amniote, Eocaptorhinus.
Unfortunately, in real fossils, there are no convenient color codes or even
detectable sutures in most of the braincase. Some of the boundaries shown in
figure are only approximate. Still, they give us a pretty good idea of how the
opisthotic is laid out with respect to the other bones of the brain case and
occiput.
Before
leaving the standard condition, it is worthwhile to take a careful look at the
appearance of the braincase In another early amniote, Eocaptorhinus.
Unfortunately, in real fossils, there are no convenient color codes or even
detectable sutures in most of the braincase. Some of the boundaries shown in
figure are only approximate. Still, they give us a pretty good idea of how the
opisthotic is laid out with respect to the other bones of the brain case and
occiput.
Note that at this level of vertebrate evolution, the stapes is a really massive element. While it is certainly possible that the stapes was connected in some way to hearing, it could not have performed the task in the same way as the columella of later diapsids. Another interesting feature is the size and complex topology of the supraoccipital. The dorsal and dorsolateral processes of the supraoccipital grab tightly onto the bones of the skull table like a three- fingered claw. The exoccipitals, opisthotics and prootics form a tight band around the braincase which is further stapled into position by the broad footplate of the stapes.
In fact, the basic structure seems so strongly constructed that one wonders why it was necessary to evolve a strong paroccipital process in addition. One might speculate that the evolutionary impetus was, as it often is, the drive to develop always stronger faster, and more flexible jaw muscles. That is, the development of large temporal fenestrae for jaw musculature would interfere with a structure which depended on strong attachment to the skull table. Furthermore, the growing jaw muscles would themselves be constrained by the massive stapes. Thus, it may have been advantageous for an organism to have a strong cross brace at the back of the skull which would leave more room for the development of jaw muscles anteriorly.
But we are getting a head of ourselves, so to speak. First we must try to determine how this distinctively amniote occipital structure evolved from its fishy forbearers. For this portion of the story, we must turn to a careful analysis of occiput evolution from Acanthostega to the Age of Bob. As with most early tetrapod problems, the inquiry gets bogged down in the swamps of the Carboniferous. But we will tread lightly and try not to get sucked in.
 Not
to belabor the obvious, but our ancestors were fish. For that reason the
earliest terrestrial vertebrates did not come equipped with an occiput, as we
understand the term. Fishes live in a relatively supportive medium. The are
quite happy with an unrestricted notochord. Consequently, they have no need for
an elaborate joint between the spine and the cranium. To be sure, teleost fishes
have a bone which is called the opisthotic. However, it is not homologous to the
tetrapod opisthotic and the teleost otic capsule is all of a piece.
Not
to belabor the obvious, but our ancestors were fish. For that reason the
earliest terrestrial vertebrates did not come equipped with an occiput, as we
understand the term. Fishes live in a relatively supportive medium. The are
quite happy with an unrestricted notochord. Consequently, they have no need for
an elaborate joint between the spine and the cranium. To be sure, teleost fishes
have a bone which is called the opisthotic. However, it is not homologous to the
tetrapod opisthotic and the teleost otic capsule is all of a piece.
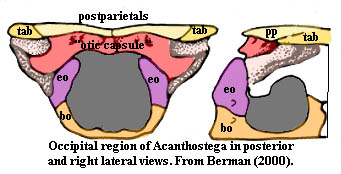
The occiput of Acanthostega, such as it is, is therefore a relatively simple affair. Apparently the exoccipitals lie directly under the otic capsule which itself forms the dorsal margin of the foramen magnum. At least this is the way it is illustrated by Berman (2000). Jarvik's interpretation of Ichthyostega is quite different. However, the present fashion is to regard Acanthostega as closest to the tetrapod stem, while Ichthyostega is relegated to the role of an atypical offshoot.
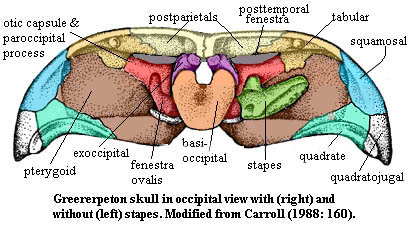 This
being the case, the interpretation given to the colosteid,
Greererpeton, may be a
little surprising. However, here there is no controversy. All seem to be agreed
that the exoccipitals rest almost on top of the otic capsule. This is important,
because Greererpeton is probably the most primitive well-known occiput
from something that is clearly not a fish. "Fish" is, of course, not a
term with much precision. Greererpeton was probably an aquatic
organism. It may well have had gills, at least in some larval form. However, it
did not have opercular bones; and its pectoral girdle was largely free of the
skull. Given that Greererpeton was as close as we can find to Acanthostega
in the direction of the amniotes, it may be legitimate to ask whether its
exoccipitals are really homologous with those of Acanthostega. But that
is a question for another day and another bone. For our purposes it is
sufficient to note that Greererpeton certainly did have a paroccipital
process of sorts. However, given its orientation and position -- a short
anterodorsal bar terminating on the tabular -- it looks more like a process of
the prootic than an occipital contribution from the opisthotic. Because the otic
capsule is not divided into separate ossifications, we can't tell whether this
structure is really a paroccipital process of the type found in other
terrestrial vertebrates. In any case, this is clearly a very primitive occiput.
This
being the case, the interpretation given to the colosteid,
Greererpeton, may be a
little surprising. However, here there is no controversy. All seem to be agreed
that the exoccipitals rest almost on top of the otic capsule. This is important,
because Greererpeton is probably the most primitive well-known occiput
from something that is clearly not a fish. "Fish" is, of course, not a
term with much precision. Greererpeton was probably an aquatic
organism. It may well have had gills, at least in some larval form. However, it
did not have opercular bones; and its pectoral girdle was largely free of the
skull. Given that Greererpeton was as close as we can find to Acanthostega
in the direction of the amniotes, it may be legitimate to ask whether its
exoccipitals are really homologous with those of Acanthostega. But that
is a question for another day and another bone. For our purposes it is
sufficient to note that Greererpeton certainly did have a paroccipital
process of sorts. However, given its orientation and position -- a short
anterodorsal bar terminating on the tabular -- it looks more like a process of
the prootic than an occipital contribution from the opisthotic. Because the otic
capsule is not divided into separate ossifications, we can't tell whether this
structure is really a paroccipital process of the type found in other
terrestrial vertebrates. In any case, this is clearly a very primitive occiput.
 Since
Greererpeton is generally felt to be closest to the temnospondyls, the
next logical step is to compare it with the temnospondyls. [2].
But here we run into something rather unexpected. Berman bases his model of the
temnospondyl occiput on Platyhystrix.
He illustrates this organism as having a broad, plate like opisthotic lateral to
the exoccipitals, with a squarish corner homologous to the paroccipital process.
Perhaps a more typical example of the temnospondyl condition is found in Luzocephalus.
Luzocephalus and many other temnospondyls have something a great deal
more like a paroccipital process which is created by a laterodorsal process of
the exoccipital sutured to a corresponding ventromedial extension of the
tabular. Other temnospondyls form a similar structure with the exoccipital and
the postparietals. At least one temnospondyl, Lapillopsis,
apparently has both structures.
Since
Greererpeton is generally felt to be closest to the temnospondyls, the
next logical step is to compare it with the temnospondyls. [2].
But here we run into something rather unexpected. Berman bases his model of the
temnospondyl occiput on Platyhystrix.
He illustrates this organism as having a broad, plate like opisthotic lateral to
the exoccipitals, with a squarish corner homologous to the paroccipital process.
Perhaps a more typical example of the temnospondyl condition is found in Luzocephalus.
Luzocephalus and many other temnospondyls have something a great deal
more like a paroccipital process which is created by a laterodorsal process of
the exoccipital sutured to a corresponding ventromedial extension of the
tabular. Other temnospondyls form a similar structure with the exoccipital and
the postparietals. At least one temnospondyl, Lapillopsis,
apparently has both structures.
Note that in Lapillopsis, the stapes articulates with a process of the "exoccipital." Furthermore, the jugular foramen -- usually at the exoccipital-opisthotic boundary -- is well inside the exoccipital. What this suggests is that the exoccipital and opisthotic are fused or, perhaps more likely, that they are the same bone. Therefore, the "paroccipital process" of Greerpeton and Platyhystrix is quite possibly an unrelated process of the prootic or the old, unitary otic capsule. Conceivably, the exoccipital of Acanthostega may also be unrelated to the temnospondyl bone of the same name or, perhaps the opisthotic is itself derived from an extension of the exoccipital onto the otic capsule instead of a subdivision of the original otic capsule. [3] This supposition may be supported by the very primitive temnospondyls Eryops and Edops, in which the exoccipitals unite above the foramen magnum and roof the cranial cavity above the otic capsule. Berman (2000: 945).
![]() This
looks orderly, but rather strange. What happens if we take a step in another
direction from Greerpeton, toward the amniotes? Possibly the closest we
can come to this is Megalocephalus,
a baphetid. The baphetids are weird, possibly alligator-like forms with
enormous keyhole orbits. Their position is much debated, but the general feeling
is that they diverged from the line leading to amniotes later than the
temnospondyls. This is an area of phylospace we have not explored much, so it is
hard to comment (undoubtedly this is for the best). At this level, something
which is clearly related to the amniote opisthotic appears on the surface and
forms a process joining with the tabular. In baphetids and anthracosaurs, the
opisthotics join medially over the top of the foramen magnum. In amniotes, this
central region is replaced by a (usually) unpaired supraoccipital formed from
the ossification of cartilaginous bands which normally join the two otic
capsules. Berman asserts that the supraoccipital is already present at this
level and is continuous with the opisthotic.
This
looks orderly, but rather strange. What happens if we take a step in another
direction from Greerpeton, toward the amniotes? Possibly the closest we
can come to this is Megalocephalus,
a baphetid. The baphetids are weird, possibly alligator-like forms with
enormous keyhole orbits. Their position is much debated, but the general feeling
is that they diverged from the line leading to amniotes later than the
temnospondyls. This is an area of phylospace we have not explored much, so it is
hard to comment (undoubtedly this is for the best). At this level, something
which is clearly related to the amniote opisthotic appears on the surface and
forms a process joining with the tabular. In baphetids and anthracosaurs, the
opisthotics join medially over the top of the foramen magnum. In amniotes, this
central region is replaced by a (usually) unpaired supraoccipital formed from
the ossification of cartilaginous bands which normally join the two otic
capsules. Berman asserts that the supraoccipital is already present at this
level and is continuous with the opisthotic.
While we have skipped over a great deal of detail, including the entire point of Berman's paper and all of the strange variations worked on these basic themes among the lepospondyls and living amphibians, this brings us more or less up to the level of Bob. It is clear only that the origin and evolution of the opisthotic is closely tied to the exoccipitals and the supraoccipital. However, the precise relationships are still vague. Note that Bob's opisthotic is more or less continuous with the exoccipitals -- an apparently primitive configuration -- and that this seems to be the normal pattern for reptiles. In some closely related basal or earliest amniotes like Diadectes and in early synapsids, the arrangement looks more like Megalocephalus, where the opisthotic and supraoccipital are more intimate. Turtles, fittingly enough, seem to begin somewhere in between, although parieasaurs have the Bob-like reptilian pattern.
 We
have spent an unusually long time trying to understand the Standard Condition,
and without much success at that. This page threatens to become so encumbered
that it resembles one of Prof.
Paul Olsen's bandwidth-busting Mesozoic life pages, sadly without the quality that
marked his work. We must now race
through the amniotes like Benni
McCarthy running past a couple of winded fullbacks.
We
have spent an unusually long time trying to understand the Standard Condition,
and without much success at that. This page threatens to become so encumbered
that it resembles one of Prof.
Paul Olsen's bandwidth-busting Mesozoic life pages, sadly without the quality that
marked his work. We must now race
through the amniotes like Benni
McCarthy running past a couple of winded fullbacks.
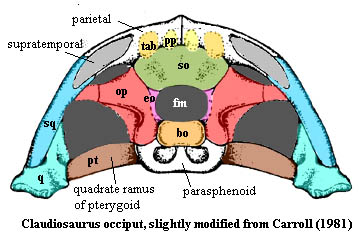
Among the reptiles, as we've seen, the opisthotic tends to consort with the exoccipitals. It has little choice in the matter, since the supraoccipital tends to retreat into austere seclusion as a vertical element dedicated to loftier, or at least more dorsal, goals such as keeping the skull table together with the occiput. This is particularly necessary because the tabular, supratemporal and postparietal bones retreat over the course of the Permian. Those vertical anchors are weakened and finally lost. Both of these developments have a liberating effect on the opisthotic, which now combines with the exoccipitals to stretch across the back of the skull to form the lateral arms of the typical "Maltese Cross" occiput of archosaurs, illustrated elsewhere. As we can see from Claudiosaurus [Carroll (1981)], this pattern was well-established in basal diapsids, before archosaurs and lepidosaurs diverged.
Each of these groups has a minor variation on the general theme. In pythonomorphs and varanoids, the crista prootica is particularly well developed and actually extends posteriorly to participate in the paroccipital process. Rieppel & Zaher (2000). In archosaurs, there is a strong tendency for the entire occiput to fuse into one inseparable mass, invariably so in theropods. Currie & Zhao (1993). However, developmentally, the pattern is the same as in all other reptiles.
 In
synapsids, this simplification does not take place. the tabulars and
postparietals are retained. The opisthotic-supraoccipital complex is likewise
retained as a massive, plate-like paroccipital element. However, as the temporal
fenestra expands to ludicrous proportions in the cynodonts, the skull table is
reduced to a saggital crest formed by the parietal. The postparietals, tabulars and
the paroccipital plate are all squeezed into a vertical triangular occipital
area at the back of the skull, framed by the squamosals. Since there is no
particular advantage to having separate ossifications in such a structure, this
development set the stage for the more or less unitary skull of the mammals [4].
In
synapsids, this simplification does not take place. the tabulars and
postparietals are retained. The opisthotic-supraoccipital complex is likewise
retained as a massive, plate-like paroccipital element. However, as the temporal
fenestra expands to ludicrous proportions in the cynodonts, the skull table is
reduced to a saggital crest formed by the parietal. The postparietals, tabulars and
the paroccipital plate are all squeezed into a vertical triangular occipital
area at the back of the skull, framed by the squamosals. Since there is no
particular advantage to having separate ossifications in such a structure, this
development set the stage for the more or less unitary skull of the mammals [4].
The opisthotic is unlike any other bone we have reviewed thus far. Its origins, relationships and evolution remain uncertain. It is nominally a chondrocranial bone of the braincase, but its most obvious function and associations are with bones of the occiput. It is closely associated, and often fused with either the exoccipitals, the supraoccipital, or even both. It may, in fact, share a common origin with one or both. Since it has no real homologues outside the "Amphibia," it ought to be , as Berman argues, a good phylogenetic marker for the tetrapod stem group; but somehow, its pattern seems to elude accurate classification.
[1] Bob is loosely based on Paleothyris from Carroll (1988: 194). However, numerous changes have been made to better illustrate the general amniote condition, and any resemblance to actual occiputs, living or dead, is purely coincidental.
[2] Platyhystrix is taken from Berman (2000), labeled in accordance with the original. Luzocephalus is from Warren (1998). The original is unlabelled. I had some difficulty with this figure, and some areas (not those of major interest) may be mislabeled or shown with misplaced borders. Lapillopsis is from Yates (1999). This is labeled per the original, except for the jugular foramen which is unlabelled in the original.
[3] Readers should not have to be reminded to take these speculations with a massive grain of salt. As always, the analysis is for amusement and exercise, and should not be confused with real science.
[4] It seems to be one of those unwritten rules of paleontology that no one illustrates a mammaliform in occipital view. To be sure, there isn't a lot to show. Nevertheless, this is somewhat frustrating for those of us who wish to illustrate the full range of vertebrate cranial morphology.
| Page Back | Unit Home | Glossary | Page Top | Page Next |
checked ATW060127