| Palaeos: | Tetrapoda | |
| The Vertebrates | Baphetidae |
| Page Back | Unit Home | Unit Dendrogram | Unit References | Taxon Index | Page Next |
| Unit Back | Vertebrates Home | Vertebrate Dendrograms | Vertebrate References | Glossary | Unit Next |
|
Abbreviated Dendrogram
SARCOPTERYGII | TETRAPODA |--Acanthostega `--+--Ichthyostega `--+--Crassigyrinus `--+--Colosteidae | `--Greererpeton `--+--Baphetidae | |--Loxomma | `--+--Baphetes | `--Megalocephalus |--Eucritta `--Tetrapoda* |--TEMNOSPONDYLI `--+--LEPOSPONDYLI `--REPTILIOMORPHA |
Contents
Overview |
 Loxomma allmanni, Visean of Scotland. Like all baphetids, this animal is known almost entirely only from fossil skulls, hence the reconstruction of the rest of the body as a typical large temnospondyl crocodile-like form is purely speculative, although not unreasonable. Life reconstruction, by Dmitry Bogdanov, Wikipedia, GNU Free Documentation/Creative Commons Attribution |
 |
| Unnamed baphetid in (more or less) dorsal view from Prof. Jennifer Clack. |
Baphetidae: (= Loxommatidae). As normally used, probably a plesion of stem tetrapods, rather than a clade. Here, we stick to the undoubted clade comprised of Baphetes, Loxomma, and Megalocephalus, consistent with [B77].
Range: Early Carboniferous (Viséan) to Late Carboniferous (Westphalian D or late Moscovian) of Europe and North America.
Phylogeny: Tetrapoda ::::: Tetrapoda* + Eucritta + *: Loxomma + (Baphetes + Megalocephalus).
Introduction: The baphetids, or loxommatids as they were formerly called [ML98], were large aquatic predators of the Carboniferous period. Not only are they included among the very early tetrapods, but they were also among the first of the Carboniferous fossil tetrapods to be found and were originally described in 1850 by William Dawson. They are known mainly from skulls, very little skeletal material. But even from this quite a lot can be deduced. The presence of lateral lines and the long rows of needle-like teeth show that they were fish-eaters. The curious keyhole-shaped orbit indicates the presence either of a gland or, it has been suggested, some sort of electrosensory organ. It may even represent an early form of skull fenestration for jaw muscles. It is not clear whether the four genera assigned to this group are really closely related.
Baphetids have been previously considered primitive temnospondyls and more recently batrachosaurs (reptile-like amphibia). It is likely however that they represent one more of a number of early Carboniferous tetrapod radiations. Computer-assisted phylogenetic analyses of a data matrix using characteristics of most of the major groups of terrestrial vertebrates place the Baphetids close to the ancestry of subsequent tetrapod groups. For reference to this, and a number of alternative and previously considered trees, see: Michel Laurin's on-line essay Phylogeny of Terrestrial Vertebrates. It seems from the various alternative trees that the baphetids were cousins of the crassigyrinids, although whether the crassigyrinids evolved from the baphetids (as the earlier cladistic trees indicate, see also Panchen ) or the baphetids from the crassigyrinids is not yet certain. (adapted from MAK 930719)
 Characters: Supposedly croc-like (post-cranial skeleton essentially
unknown until very recently); shallow skull; skull dermal bones similar to Edopoidea
[B77]; septomaxilla appears as ornamented,
relatively large surface bone [C02]; lateral line grooves present (but somewhat reduced); keyhole-shaped orbits formed by excavation of the lacrimal and
prefrontal anterior to the orbit (see figure)
and also bordered by jugal [ML98]; antorbital excavation at least as large as
orbit [ML98]; strongly embayed spiracular notch, notch
at posterolateral border of skull table, opening posteriorly [B77]; small
tabulars with posterior process for attachment of mandibular depressors [B77]; orbits at mid-length of skull [B77]; orbits (sensu stricto) circular
[B77]; cheek & skull roof sutured; quadrate
well behind occiput (as most early tetrapods) [B77]; palatoquadrate
probably still present as a unit, although partly formed in cartilage [B77];
exoccipitals do not reach postparietals or continue under skull table (compare Edops)
[B77]; supraoccipital absent (not known for all genera) [B77], contra
[B00]; stapes proximally broad; palate closed;
braincase heavily ossified and extensively attached to skull roofing bones
[B77]; opisthotic with two pairs of exoccipital facets (see Megalocephalus)
[B77]; nerves IX, X, XI exit between exoccipital facets [B77]; otic capsule with
two short, stout columnar processes articulating with exoccipital columns, anterior of which articulates with parietal and
supratemporal [B77]; premaxilla form stout buttresses anteriorly on palate [B77];
adsymphysial plate present [ML98]; teeth
"labyrinthodont" [C02]; fangs round at base, becoming laterally
compressed at crown, with mesial & distal keels [ML98]; parasymphysial plate
with at most 2 teeth [ML98]; coronoids
have no teeth or denticles; double tooth row on dentary; shows some tendency to
consolidation of jaw.
Characters: Supposedly croc-like (post-cranial skeleton essentially
unknown until very recently); shallow skull; skull dermal bones similar to Edopoidea
[B77]; septomaxilla appears as ornamented,
relatively large surface bone [C02]; lateral line grooves present (but somewhat reduced); keyhole-shaped orbits formed by excavation of the lacrimal and
prefrontal anterior to the orbit (see figure)
and also bordered by jugal [ML98]; antorbital excavation at least as large as
orbit [ML98]; strongly embayed spiracular notch, notch
at posterolateral border of skull table, opening posteriorly [B77]; small
tabulars with posterior process for attachment of mandibular depressors [B77]; orbits at mid-length of skull [B77]; orbits (sensu stricto) circular
[B77]; cheek & skull roof sutured; quadrate
well behind occiput (as most early tetrapods) [B77]; palatoquadrate
probably still present as a unit, although partly formed in cartilage [B77];
exoccipitals do not reach postparietals or continue under skull table (compare Edops)
[B77]; supraoccipital absent (not known for all genera) [B77], contra
[B00]; stapes proximally broad; palate closed;
braincase heavily ossified and extensively attached to skull roofing bones
[B77]; opisthotic with two pairs of exoccipital facets (see Megalocephalus)
[B77]; nerves IX, X, XI exit between exoccipital facets [B77]; otic capsule with
two short, stout columnar processes articulating with exoccipital columns, anterior of which articulates with parietal and
supratemporal [B77]; premaxilla form stout buttresses anteriorly on palate [B77];
adsymphysial plate present [ML98]; teeth
"labyrinthodont" [C02]; fangs round at base, becoming laterally
compressed at crown, with mesial & distal keels [ML98]; parasymphysial plate
with at most 2 teeth [ML98]; coronoids
have no teeth or denticles; double tooth row on dentary; shows some tendency to
consolidation of jaw.
Links: Loxommatidae (Baphetidae); Air Breathers of the Coal Period; TREE2000.pdf; Eucritta; AE TREE2000.pdf; Report of Activities 1999: Calder No. 1; Class:Amphibia; Amphibian Skull.
Notes: [1] With the reinterpretation of Ichthyostega and its relatives as aquatic forms, baphetids are good candidates for the spot of first amphibious tetrapod group. If so, baphetids may be a rather important taxon. Both Clack's discovery of Eucritta and recent phylogenetic studies (Ahlberg & Clack (1998)) could support this conclusion. [2] The current candidates for the function of the preorbital excavation are (a) a salt gland, (b) an electrosensory organ; and (c) the pterygoideus muscle. Of the three, the third seems most consistent with the general observation that most serious cranial re-engineering is related to jaw mechanics. This weird adaptation may simply be an early, and very different, type of skull fenestration.
References: Ahlberg & Clack (1998); Beaumont (1977) [B77]; Berman (2000) [B00]; Clack (2002) [C02]; Milner & Lindsay (1988) [ML98]. ATW020820.
 Loxomma:
L. allmani (almanni?) Huxley 1862; L. acutirhinus, L. rankini
Loxomma:
L. allmani (almanni?) Huxley 1862; L. acutirhinus, L. rankini
Range: Early Carboniferous (Viséan?!) to Late Carboniferous (Bashkirian, Westphalian A) of Europe (Britain).
Phylogeny: Baphetidae : (Baphetes + Megalocephalus) + *.
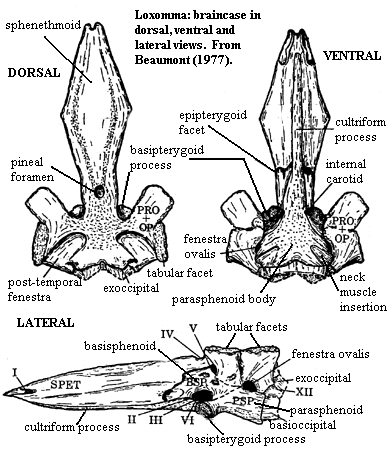
Characters: dermal skull: skull ~25 cm and flattened [C02], contra [B77]; snout bluntly rounded; premaxilla long [B77]; lacrimal reaches nares [B77]; nares circular, at anterolateral margin of skull [B77]; septomaxilla not known [B77]; antorbital vacuity present and about same size as orbit [B77]; margins of vacuity solid, formed by lacrimal and nares "united" with buttresses from palatal bones, forming continuous, smooth-sided wall [B77]; orbits high on skull [C02]; orbits roughly at mid-skull length [B77]; intertemporal present [B77] [C02]; skull table large and square [B77]; pineal opening on slight raised boss [B77]; maxilla reaches quadratojugal, excluding jugal from skull margin [B77]; strong embayment between skull table & cheek area = probable spiracular notch [C02]; occiput: occipital arches weakly integrated into skull [B77]; exoccipital with limited role in braincase (primitive) [B77]; extended opisthotic forms paroccipital process [B77]; paroccipital process articulates only with tabular of skull table [B77]; opisthotics fused on the occiput? [B77]; braincase: (see image under Baphetidae) braincase strongly attached to dermal roofing bones [B77]; otic capsule with two strong lateral processes, each with exoccipital facet [B77]; otic capsule posterolateral wall from more posterior (opisthotic) process [B77]; same process forms dorsal border of fenestra ovalis and also floors fossa inside posttemporal fenestra, articulates with tabular dorsally (possible $) [B77]; floor of fenestra ovalis from parasphenoid [B77]; more anterior (prootic) process articulates with supratemporal & parietal [B77]; V-shaped depressions on ventral face of parasphenoid for attachment of neck muscles [B77]; basisphenoid ascends laterally anterior to otic capsule with facets for epipterygoid [B77]; sphenethmoid ossified and extends to presumed position of nasal capsules [B77]; sphenethmoid strongly ossified as interorbital septum [B77]; palate: palate not well known [B77]; interpterygoid vacuities absent [B77]; palatal anterior vacuity absent [B77]; premaxilla sutures with vomer along entire length of vomer [B77]; dentition: premaxilla with 8 teeth [B77]; premaxillary alveolar shelf broadens & teeth larger distally [B77]; jaws & palate with large, recurved teeth with slight keels [C02]; dentary parasymphysial tusks probably short [B77]; maxilla marginal teeth and alveolar shelf larger distally [B77]; maxillary marginal tooth count <30 [B77]; vomer, palatine & ectopterygoid each with tusk pair on a raised boss [B77]; palatine palatal ramus with shagreen of denticles [B77]; coronoids and prearticular with slight shagreen of denticles [AC98]; dermal pit & ridge ornament poorly developed in some specimens [B77].
Links: Class:Amphibia.
References: Ahlberg & Clack (1998) [AC98]; Beaumont (1977) [B77]; Clack (2002) [C02]. ATW020820.
| Page Back | Unit Home | Page Top | Page Next |
checked ATW040813