
Life restoration of Labidosaurus hamatus
| Eureptilia | ||
| The Vertebrates | Captorhinidae (3) |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abreviated Dendrogram
Amniota
├─Synapsida
└─┬─Anapsida
│
└─Eureptilia
├─Coelostegus
└─┬─┬─Thuringothyris
│ └─Captorhinidae
│ ├─Concordia
│ └─┬─Romeria
│ └─┬─Captorhinus
│ └─┬─Captorhinikos
│ └─┬─Labidosaurus
│ └─Moradisaurinae
└─Romeriida
├─Paleothyris
└─┬─Protorothyrididae
└─Diapsida
|
Contents
Overview |
|
| Life restoration of Labidosaurus hamatus |
Captorhinidae : Concordia + (Romeria + (Protocaptorhinus + (Rhiodenticulatus + (Saurorictus + (Captorhinikos + (Labidosaurus + Moradisaurinae))))) + * ))
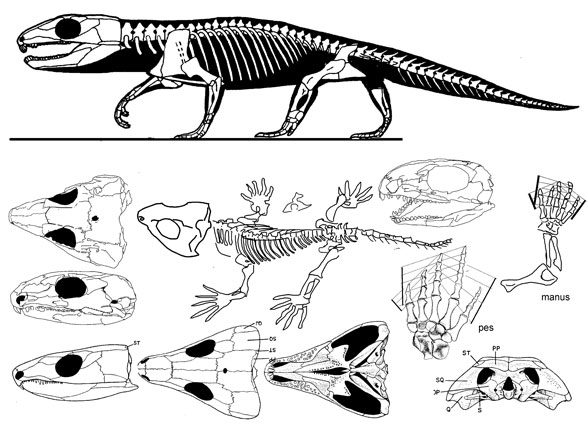 |
| Captorhinus aguti (Cope 1882) Artinskian, Early Permian, ~40 cm in length, studies of skull and skeleton. Illustration copyright David Peters, reproduced with permission |
Captorhinus is the type genus of the family Captorhinidae. This genus comprises the single-tooth-rowed species C. laticeps and C. magnus as well as the multiple-tooth-rowed C. aguti. All members of the genus are exclusively known from Lower Permian terrestrial deposits of Texas and Oklahoma.
Scientific History: Captorhinus is the longest-known genus of the family and its scientific record dates back to the 1880s. In 1882 E. D. Cope described Ectocynodon aguti and later assigned it to his genus Pariotichus leading to the new combination P. aguti. In 1909 S. W. Williston described the species P. laticeps.
A first revision of these "cotylosaurs" by E. C. Case in 1911 revealed that most of the representatives of Pariotichus belong to another genus erected by Cope in 1895: Captorhinus. Case identified four species of Captorhinus: C. aguti, C. angusticeps, C. isolomus, and C. aduncus. In that course P. laticeps was synonymized with C. isolomus. Moreover Case de facto declared Ectocynodon a nomen dubium due to the bad state of preservation of its type material. Strictly speaking the state of preservation of the holotype of Captorhinus is not much better, however, Case deemed it diagnostic at that time. Seen that Ectocynodon is thus the senior synonym proper it provides a good example for a nomen oblitum, because it has never been used in captorhinid taxonomy since 1911.
In 1959 a second revision of the captorhinid fossils by R. J. Seltin lead to the recognition of only one species of Captorhinus: C. aguti. Oddly Seltin (1959) erected the new species Labidosaurus oklahomensis on the basis of a small single-tooth-rowed skull with Captorhinus-like appearance. Later doubts were casted on the assignment of the material formerly referred to P. laticeps to C. aguti because of the presence of a single tooth row, unlike to all other remains of C. aguti which show multiple tooth rows in at least one branch of the lower or upper jaw. In 1979 M. J. Heaton thus erected the genus Eocaptorhinus for these exclusively single-tooth-rowed remains leading to the new combination E. laticeps. In addition Heaton assigned Seltin's L. oklahomensis to this species. Finally E. laticeps was reassigned to Captorhinus by several authors because of its general similarity to C. aguti (see below), but this time retained its status as distinct species.
In 2001 the scientifically youngest species of Captorhinus was reported by Kissel et al. and subsequently, in 2002, was given the name C. magnus.
Characteristics of the genus: Captorhinus is characterized by a series of cranial traits that occur in other basal captorhinids as well, e.g. relatively large eye openings, shagreen dentiton on palatal bones, a distinctive sculptural pattern on the bones of the dermal skull roof and on the bones of the external surface of the mandible, and a down-curved, hook-like premaxilla. However within the genus one species existst that has multiple tooth rows, a feature not known from other basal captorhinids, such as Romeria or Protocaptorhinus. In addition the posterior teeth of the maxillary are not simple cones as in more basal captorhinids but are either chisel-shaped or ogival. Also the posterior part of the skull is clearly more expanded laterally, being reminiscent rather to the moradisaurines than to the basal members of the family. A unique feature of Captorhinus may be the retroarticular process of the lower jaw being conspiciously longer than broad, since this is as yet not seen in any other captorhinid taxon.
Type species: C. aguti is the type species of the genus. It is without doubt the by far best known captorhinid species. Hundreds of well preserved, articulated skulls and skull fragments are collected at the famous Dolese Brothers quarry near Fort Sill in Oklahoma.
C. aguti is distinguished primarily from C. laticeps and C. magnus in having multiple rows of teeth on the maxillary and dentary bones. These rows are, in contrast to the tooth rows seen in moradisaurines, not oriented parallel to the lateral margins of the jaws but are oriented obliquely to that margins, in a way that each row is followed posterolaterally by the next one. Thus, up to eight rows are observed in C aguti without the dentigerous bones forming the strongly medially expanded tooth plates which characterize the jaws of the moradisaurines. Nevertheless the toothed areas of maxillary and dentary are broader mediolaterally than in the single-tooth-rowed captorhinids.
Second species: C. laticeps is distinguished form C. aguti in lacking multiple tooth rows in all four branches of the jaws. In almost all other respects it is very similar to C. aguti. C. laticeps differs from C. magnus in being considerably smaller and in having chisel-shaped instead of ogival teeth on the posterior portion of maxillary and dentary.
Synonymy of Eocaptorhinus: Eocaptorhinus is considered a synonym of Captorhinus. The genus was erected by Malcolm J. Heaton in 1979 including fossils hitherto referred to Pariotichus laticeps Williston leading to the new combination Eocaptorhinus laticeps. Heaton chose this name meaning "early Captorhinus because the remains of this animal are striking similar to Captorhinus aguti, with the exception of lacking the characteristic multiple tooth rows, but come from an geologically older horizon. Disregarding the principles of cladistics Heaton made an stratophenetic approach in elucidating the evolution of captorhinids, postulating an anagenetic Romeria -> Protocaptorhinus -> Eocaptorhinus -> Captorhinus model.
More recent examinations of Eocaptorhinus and Captorhinus aguti revealed that they share such a great number of non-dentitional traits that distinction on a generic level became obsolete. Moreover in one locality (Ft. Sill, Oklahoma) captorhinid remains with both, single tooth rows and multiple tooth rows are present without any possibility of stratigraphic differentiation of the fossils found at that time. In addition it has been shown that in C. aguti some specimen have a single tooth row on one and a double tooth row on the other branch of the jaw. Thus, despite minor differences in postcranial body structure, it has been ambiguous wether C. laticeps and C. aguti are truly distinct species and if not in which way both forms are related: as sympatric subspecies, as males and females, or afterall as distinct ontogenetic stages of one single species.
Then a significant difference in the shape of the teeth of both forms was recognized: C. aguti was discovered to have teeth that resemble rather a pointed arch (also called ogive) whereas C. laticeps has teeth that are rather chisel-shaped. However this seemed not sufficient to legitimate the "two-species-concept".
The discovery of a new species, Captorhinus magnus, throwed light on this obscure relationship in a cladistical way. C. magnus is approximately as twice as large as C. aguti and its jaws are equipped with only one single row of, however ogival, teeth. Most important is the fact that C. magnus is unequivocally distinct from C. aguti in having an concave articulation surface at the distal end of the femur whereas in C. aguti this articulation surface is convex, a feature hardly relatable to ontogeny. Sexual dimorphism could also be excluded because further collections at the Ft. Sill site obtained that specimen of C. magnus are more abundant in the lower parts of the section whereas C. aguti is more abundant in the higher part. In case of sexual dimorphism a by far more regular distribution would be expected. Finally, inclusion of the new taxon in the phylogenetic analyses yielded that C. laticeps and C. aguti are no longer sister clades but C. aguti is sister to C. magnus. Thus, any conspecific or closer relationship of C. laticeps and C. aguti can be ruled out due to the close relationship of C. aguti to another, species of Captorhinus that is clearly not conspecific with C. aguti.
Third species: C. magnus differs from C. aguti in having only one single row of teeth on dentary and maxillary. From C. laticeps it is distinct in being twice as large and in having ogival teeth in the posterior portion of maxillary and dentary. Moreover the distal articulation surface of the femur is concave whereas in C. aguti and C. laticeps it is convex.
Phylogeny: Captorhinus comprises rather derived representatives of basal captorhinids. The dentition, accompanied with the laterally widened posterior portion of the skull, shows a certain degree of specialization compared to the more basal forms.
Below follow succinct accounts on the recognized species of Captorhinus
BROOM, R. (1910): A comparison of the Permian reptiles of North America with those of South Africa. Bulletin of the American Museum of Natural History 28: 197-234
COPE, E.D. (1895): The Reptilian Order Cotylosauria. Proceedings of the American Philosophical Society 34 (149): 436-457
DODICK, J.T. and MODESTO, S.P. (1995): The Cranial Anatomy of the Captorhinid Reptile Labidosaurikos meachami from the Lower Permian of Oklahoma. Palaeontology 38 (3): 687-711
HEATON, M.J. (1979): Cranial Anatomy of Primitive Captorhinid Reptiles from the Late Pennsylvanian and Early Permian, Oklahoma and Texas. Oklahoma Geological Survey, Bulletin vol. 127. The University of Oklahoma, Norman, 83 p.
KISSEL, R.A., DILKES, D.W. and REISZ, R.R. (2002): Captorhinus magnus, a new captorhinid (Amniota: Eureptilia) from the Lower Permian of Oklahoma, with new evidence on the homology of the astragalus. Canadian Journal of Earth Science, 39 (9): 1363-1372
MODESTO, S.P. (1998): New information on the skull of the Early Permian reptile Captorhinus aguti. PaleoBios 18 (2/3): 21-35
SELTIN, R.J. (1959): A review of the family Captorhinidae. Fieldiana Geology 10 (34):461-509
OLSON, E.C. (1962a): Late Permian Terrestrial Vertebrates, U.S.A. and U.S.S.R. Transactions of the Philosophical Society of America, New Series 52 (2): 1-224
OLSON, E.C. (1962b): Part II. - The Osteology of Captorhinikos chozaensis Olson. pp. 49-68 in: Permian Vertebrates from Oklahoma and Texas. Oklahoma Geological Survey, Circular vol. 59. The University of Oklahoma, Norman
Weblinks: | photographs of specimen of Captorhinus aguti from the Richards Spur (Ft. Sill) locality
| photographs of specimen of Captorhinus magnus from the Richards Spur (Ft. Sill) locality
Zidane 2 June 2008
Captorhinus aguti Cope 1882
Generic name means "capture-nose", refers to the assumption that the hook-like premaxilla was used to capture prey animals. Specific epithet is derived from the extant rodent agouti (Dasyprocta) since Cope had the impression that the skull on which he based the species Ectocynodon aguti (see synonyms list) would be similar in shape to the skull of this animal
Ectocynodon aguti Cope 1882, Ectocynodon incisivus Cope 1888, Pariotichus aguti Cope 1882 (Cope 1895, new comb.), Pariotichus incisivus Cope 1882 (Cope 1895, new comb.), Captorhinus angusticeps Cope 1895, Pariotichus isolomus Cope 1895, Pariotichus angusticeps Cope 1895 (Broom 1910, ? lapsus calami), Captorhinus isolomus Cope 1895 (Case 1911, new comb.)
Family: Captorhinidae
Captorhinus laticeps Williston 1909
Generic name means "capture-nose", refers to the assumption that the hook-like premaxilla was used to capture prey animals. Specific epithet means "broad-headed"
Pariotichus laticeps Williston 1909, Captorhinus isolomus Cope 1895 (Case 1911, new comb.), Captorhinus aguti Cope 1895 (Case 1911, new comb.) (Seltin, 1959), Labidosaurus oklahomensis Seltin 1959, Labidosaurus stovalli (Olson, 1962a, b), Eocaptorhinus laticeps Williston 1909 (Heaton 1979, new comb.)
Clyde Formation, Admiral Formation, Belle Plains Formation (all Wichita Group), Wellington Formation (Sumner Group)
Lower Permian : Leonardian
Remains found in Baylor, Wilbarger, and Wichita Counties, Texas, USA and in Kay County, Oklahoma, USA. Inhabited "Eastern Shelf" of Midland Basin, Anadarko Basin, northwestern Pangaea
Captorhinus magnus Kissel et al. 2002
Generic name means "capture-nose", refers to the assumption that the hook-like premaxilla was used to capture prey animals. Specific epithet means "large"
Captorhinidae ::: Captorhinus + ((Labidosaurus + Moradisaurinae) + *)
Upper Vale Formation
Kungurian
Texas
E. C. Olson. 1954. Fauna of the Vale and Choza: 9. Captorhinomorpha. Fieldiana: Geology 10(19):211-218
Labidosaurus hamatus Cope 1895
Captorhinidae ::: Captorhinus + (Captorhinikos + (Moradisaurinae + *))
 |
| Drawing of a skull of Labidosaurus hamatus (modified from Case, 1911) |
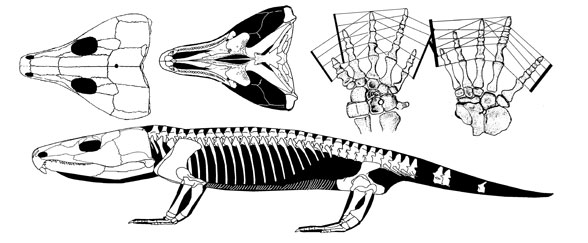 |
| Labidosaurus hamatus Cope 1895 Artinskian, Early Permian, ~75 cm in length, studies of skull, hand, and skeleton. Illustration copyright David Peters, reproduced with permission |
Generic name means "lipped reptile". Specific epithet means "hooked"
Pariotichus hamatus Cope 1895, Labidosaurus broilii Case 1911
Family: Captorhinidae
deepest part of Clear Fork Group (former "Arroyo Formation")
Lower Permian : Leonardian
Remains found in Baylor County, Texas, USA. Inhabited "Eastern Shelf" of Midland Basin, northwestern Pangaea
>Labidosaurus hamatus is the largest among the single-rowed captorhinids and is known from a few skulls and numerous postcranial remains coming from the Lower Permian red beds of North Texas. It is currently the only recognized species of the genus Labidosaurus
Its skull measures about 20 cm in length. Given that the trunk length in Captorhinus is approximately 3.6-fold the skull length, the overall body length in L. hamatus was more than 90 cm. Another particularity concerning the skull is the large hook-like tip of the snout, formed by the premaxillary bone. Most of the captorhinids have such a down-curved premaxillary but in L. hamatus this feature is much more pronounced. Use and function of this toothed "beak" is as yet unclear. Some authors suggested it served as a tool for digging, but absence of characteristic scratch marks on the anterior teeth does not support this hypothesis.
The skull morphology places Labidosaurus close to the large multiple tooth-rowed captorhinids, the moradisaurines. Because herbivory is assumed for the latter the diet of L. hamatus may have included plants but its single rows of sharp conical teeth, rather suitable for piercing animal prey, suggests that it was most likely an omnivore.
The vertebral column of L. hamatus consists of 25 trunk vertebrae and at least 33 caudal vertebrae. There are two temporally and spatially coexisting morphotypes present in the vertebral column of Labidosaurus: one that has vertebrae with short neural spines and long such spines alternating and another that has exclusively vertebrae with short neural spines.
L. hamatus is one of the historically longest-known members of the family and was described as early as 1895 by one of the trailblazers of US-american vertebrate paleontology Edward D. Cope (albeit under a different name of genus). Some captorhinid fossils formerly referred to Labidosaurus are today assigned either to other captorhinid genera or to L. hamatus.
COPE, E.D. (1895): The Reptilian Order Cotylosauria. Proc. Am. Phil. Soc., 34(149), pp. 436-457
COPE, E.D. (1896): Second contribution to the history of the Cotylosauria. Proc. Am. Phil. Soc., 35(151), pp. 122-139
MODESTO, S.P., SCOTT, D.M., BERMAN, D.S., MÜLLER, J. and REISZ, R.R. (2007): The skull and the paleoecological significance of Labidosaurus hamatus, a captorhinid reptile from the Lower Permian of Texas. Zool. J. Linn. Soc., 149(2), pp. 237-262
SUMIDA, S.S. (1987): Two Different Vertebral Forms in the Axial Column of Labidosaurus (Captorhinomorpha: Captorhinidae). J. Paleontol., 61(1), pp. 155-167
Zidane 25 May 2008
content Zidane; page MAK120316