Stylophora
Like a Tee-to-tum They Twirled Him
 The Stylophora were a not-overly-diverse but long-lived group of animals (Middle Cambrian to Middle Devonian - Ubaghs, 1967) that have probably received the most attention of the stem echinoderms, largely because of Jefferies' e.g. 1986) suggestion that they were ancestral to the chordates according to his "calcichordate" theory. Jefferies' theory was strongly dependent on his interpretation of the fossil homalozoans, which he referred to as "calcichordates." On this page, we take the more orthodox view that homalozoans are stem echinoderms, as suggested by their possession of a calcite skeleton with a similar ultrastructure to modern echinoderms. Jefferies held this skeleton to be ancestral for Chordata, and subsequently independently lost in each of the three chordate subphyla. However, this seems to require a lot of convergence, expecially if, as molecular phylogenies suggest, the hemichordates are actually closer to echinoderms. A stem echinoderm position for homalozoans seems much more parsimonious.
The Stylophora were a not-overly-diverse but long-lived group of animals (Middle Cambrian to Middle Devonian - Ubaghs, 1967) that have probably received the most attention of the stem echinoderms, largely because of Jefferies' e.g. 1986) suggestion that they were ancestral to the chordates according to his "calcichordate" theory. Jefferies' theory was strongly dependent on his interpretation of the fossil homalozoans, which he referred to as "calcichordates." On this page, we take the more orthodox view that homalozoans are stem echinoderms, as suggested by their possession of a calcite skeleton with a similar ultrastructure to modern echinoderms. Jefferies held this skeleton to be ancestral for Chordata, and subsequently independently lost in each of the three chordate subphyla. However, this seems to require a lot of convergence, expecially if, as molecular phylogenies suggest, the hemichordates are actually closer to echinoderms. A stem echinoderm position for homalozoans seems much more parsimonious.
Stylophora are generally united with the following three taxa - Soluta, Cincta and Ctenocystoidea - into the Homalozoa, regarded as a subphylum of Echinodermata. However, for reasons that will be covered later, this taxon appears to be paraphyletic. The four classes have little to unite them other than possession of a flattened theca and absence of pentaradial symmetry. For our purposes, "homalozoans" are regarded as a grade only, and the Echinodermata refers to the crown group of living echinoderms.
Stylophora are divided between two orders, the strongly asymmetrical Cornuta and the more bilateral Mitrata. Some authors regard certain Cornuta as forming a stem group to Mitrata, and move them over to join mitrates in an order Ankyroida. See results and discussion in Ruta 2003). They possess a theca which is always asymmetrical to some degree - even in those species where the external outline is symmetrical, the pattern of the plates that make it up is not. The broadest edge of the theca bears a plated appendage, and there is a major orifice on the edge opposite the appendage. The theca has two faces, one flat or slightly concave, one convex. In Cornuta there are often a number of openings on the convex surface to the left and below the appendage if the convex side is faced upwards and the appendage is pointed upwards - either a single lamellate structure or a series of pores. There are fewer pores on both faces of Mitrata, placed bilaterally. The appendage is divisible into three parts - a broad proximal section where it joins the theca then a median section tapering to narrower distal section. A canal runs through the centre of the appendage, with plates covering it.
 So far these are all uncontroversial features, but they are also pretty meaningless. A critical look at the previous paragraph will soon reveal that we haven't assigned a function to a single structure. Unfortunately once you start asking such questions, things start getting difficult. Almost every possible answer has been suggested at some point. To simplify matters, most reconstructions can be allied with one of two viewpoints - the "echinoderm" reconstruction and the "calcichordate" reconstruction.
So far these are all uncontroversial features, but they are also pretty meaningless. A critical look at the previous paragraph will soon reveal that we haven't assigned a function to a single structure. Unfortunately once you start asking such questions, things start getting difficult. Almost every possible answer has been suggested at some point. To simplify matters, most reconstructions can be allied with one of two viewpoints - the "echinoderm" reconstruction and the "calcichordate" reconstruction.
In the "echinoderm" reconstruction, the theca lies with the convex side upwards and the appendage is interpreted as an anterior feeding arm "aulacophore"). The central canal is then an ambulacrum with the plates over it able to open to expose the tube feet. The major orifice opposite the aulacophore is the anus, with the mouth somewhere near the ambulacrum. Ubaghs (1967) assumed it to be on the theca near the base, while Lefebvre 2003) placed it in the median section of the aulacophore). Various interpretations are given for the major openings on the theca as gonopores, hydropores and/or respiratory openings.
The "calcichordate" hypothesis, in contrast, reverses the orientation. The appendage is regarded as a tail, with the central canal probably containing a notochord. The major orifice is the mouth, with the anus near the tail. The minor openings in Cornuta represent the gill slits or pores, while those of Mitrata are more contentious. Dominguez et al. 2002) identified bilateral internal tubes in Jaekelocarpus (Mitrata) as paired gill slits, implying a secondary regain of bilateral symmetry in organ systems in this clade. While the Cornuta were interpreted as lying with the flat side ventrally, Jefferies suggested that in Mitrata the flat side was dorsal and the convex side ventral, while the tail was curved underneath to provide forward thrust. Support for this was seen in the common preservation of Mitrata with the tail flexed over the theca. As part 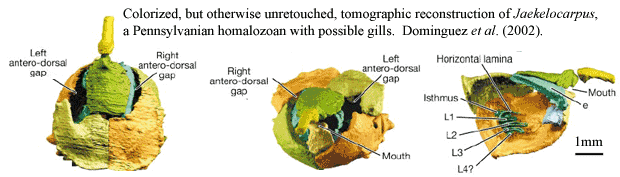 of his explanation for how this inverted orientation arose, Jefferies suggested that the mitrate tail was not entirely homologous with the cornute tail, despite its similar structure, but actually with only the proximal section. At some point, he suggested, the mitrate ancestor had lost the median and distal sections of the tail, and had then redeveloped similar structures to regain a tripartite tail.
of his explanation for how this inverted orientation arose, Jefferies suggested that the mitrate tail was not entirely homologous with the cornute tail, despite its similar structure, but actually with only the proximal section. At some point, he suggested, the mitrate ancestor had lost the median and distal sections of the tail, and had then redeveloped similar structures to regain a tripartite tail.
The "calcichordate" reconstruction is favoured here - as mentioned earlier, it fits better with the circumstantial evidence supplied by the Vetulocystidae. However, I am more sceptical of Jefferies' inverted orientation for Mitrata. For instance, Jefferies' reconstruction places the mouth ventrally while the more standard orientation would place it dorsally as in most other stem and basal echinoderms. Also his interpretation of tail homologies seems rather ad hoc. In situ fossils, it should be noted, are uniformative on this matter as both orientations are commonly found. Lefebvre (2003).
 Note that accepting Jefferies' reconstruction of Stylophora does not require us to accept his suggested phylogenetic position for them as a paraphyletic stem to Chordata with the modern chordate subphyla arising independently from various subclades. Requiring as it does the independent loss of the stereom in at least three lineages, as well as probably independent extension of a somitic structure anteriorly in Cephalochordata and Craniata, this does not seem the most parsimonious option. It seems more likely that Stylophora represent members of the echinoderm stem branching off after the origin of the stereom but before the origin of ambulacra.
Note that accepting Jefferies' reconstruction of Stylophora does not require us to accept his suggested phylogenetic position for them as a paraphyletic stem to Chordata with the modern chordate subphyla arising independently from various subclades. Requiring as it does the independent loss of the stereom in at least three lineages, as well as probably independent extension of a somitic structure anteriorly in Cephalochordata and Craniata, this does not seem the most parsimonious option. It seems more likely that Stylophora represent members of the echinoderm stem branching off after the origin of the stereom but before the origin of ambulacra.
The palaeoecology of Stylophora is nearly as contentious as its anatomy. Under the "calcichordate" model, the tail is generally assumed to be a propulsive organ, and many adherents of the "echinoderm" model also assume a locomotive function as well as a feeding one for the aulacophore. Lefebvre (2003), using the "echinoderm" model, doubted whether the aulacophore could function as such, claiming that it lacked the required flexibility and was too slender compared to the massive theca. A single slab has been found from the Devonian Hunsruck Slate of Germany which preserves specimens of Rhenocystis Mitrata) in association with infaunal trails and has been cited as evidence for a vagile lifestyle (Sutcliffe et al., 2000), but debate continues over whether the association is valid or an artifact of preservation or preparation (Lefebvre, 2003). Some Stylophora, such as the strongly asymmetrical mitrate Diamphidiocystis which has the tail displaced to the right hand side of the theca and a long scimitar-shaped articulated spine coming 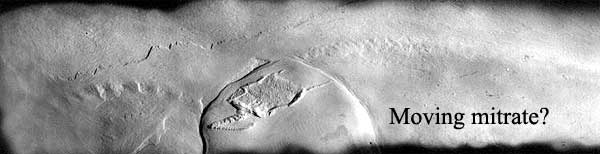 from the posterior left corner of the theca and curved towards the tail, have morphologies that would appear inconsistent with a high degree of mobility.
from the posterior left corner of the theca and curved towards the tail, have morphologies that would appear inconsistent with a high degree of mobility.
Jefferies suggested that juveniles were attached to the sediment by the ends of their tails, detaching later to become free adults. The number and arrangement of the plates seems to have become fixed early in development, with the animal growing by enlargement of pre-existing plates.
© Christopher Taylor CT050908.
Images: sources as stated. Cothurnocystis image from GeoTools.com. "Moving mitrate" image of Rhenocystis from Ferienland Hunsrück online.
page uploaded ATW050908