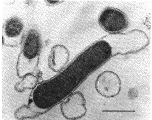
Thermotoga maritima
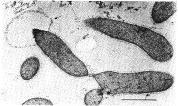
Fervidobacterium islandicum

Thermosipho sp.
| Bacteria | ||
| Bacteria | Thermotogales |
| Eubacteria | Kingdoms | |||
| Tellurobiota | Archaea |
| LUCA
├─Eubacteria
│ ├─Actinobacteria
│ └─┬─Firmicutes
│ └─Didermata
│ ├─Cyanobacteria
│ └─┬─Sphingobacteria
│ └─Proteobacteria
└─Neomura
├─Archaea
└─Eukarya
├─Chlorobionta
└─┬─Fungi
└─Metazoa
├─Deuterostomia
└─Protostomia
|
Lists |
|
|
|
| |
|
Thermotogales is a relatively minor branch of the Eubacteria consisting of only three genera: Thermotoga, Thermosipho, and Fervidobacterium. Other genera which may fall into this group include Caldotoga, Geotoga, Petrotoga, Thermopallium, and Marinotoga. All of these organisms are extremophiles. Fervidobacterium grows at relatively moderate temperatures. The other members of the group are all hyperthermophiles who flourish at 60–90° C.
The "toga" motif in the genus names derives from an unusually prominent cell envelope which can be seen most clearly in the image of Thermotoga on the right. This "toga" is apparently a reasonably ordinary outer membrane of a Gram-negative-type cell envelope.
The Thermotogales make their living by scavenging biomolecules, including amino acids, glucose, sucrose, starch, cellulose, and xylan. They are found in marine geothermal vents and non-marine hot springs, both at very high temperature but moderate pH and salinity. The Thermotogales use sulfur, rather than oxygen as an electron acceptor in metabolism, thus generating hydrogen sulfide (H2S) rather than water, as a by-product of metabolism. If the available sulfur is organically bound, considerable hydrogen gas may be produced by some species. Thermotogales are anaerobic, but can tolerate oxygen to some degree. Their ability to generate hydrogen from organic waste products, while tolerating oxygen, has encouraged some interest in the commercial use of these bacteria. van Ooteghem et al. 2001).
 The environment in which these bacteria live thus bears an unfortunate resemblance to some classic descriptions of the Underworld:
The environment in which these bacteria live thus bears an unfortunate resemblance to some classic descriptions of the Underworld:
The dismal Situation waste and wilde,
A Dungeon horrible, on all sides round
As one great Furnace flam'd, yet from those flames
No light, but rather darkness visible
Serv'd only to discover sights of woe,
Regions of sorrow, doleful shades, where peace
And rest can never dwell, hope ne'er comes
That comes to all; but torture without end
Still urges, and a fiery Deluge, fed
With ever-burning Sulphur unconsum'd
Such place Eternal Justice had prepar'd
For those rebellious, here their Prison ordain'd
In utter darkness, and their portion set
As far remov'd from God and light of Heav'n
As from the Center thrice to th' utmost Pole.
John Milton (1667), Paradise Lost, 1: 60–69.
What does an organism have to do to live on scraps, at near the boiling point of water, with little or no light or air, and in the continuous presence of active and toxic sulphur compounds? The following are some of the problems, and the strategies adopted by Thermotogales and other hyperthermophiles.
At high temperature, fat melts; and this is as true of phospholipid membranes as of lard or butter. Some fluidity is good, but it has to be balanced by the necessity for maintaining the important difference between inside and outside. (Recall, for example, the ion gradients discussed in connection with the Eubacteria). Thermotogales incorporate unusual high melting-point lipids in the cell membrane. They also make use of unusual long-chain dicarboxylic fatty acids in their membrane lipids. One possible function of these molecules is to create a tightly bound polar layer within the membrane.
At higher temperature, DNA denatures. That is, the two strands of the double helix fall apart. Hyperthermophiles use a mixture of three strategies to stabilize their DNA against thermal denaturation. (1) the amount of free water in the cell is minimized. DNA can melt because water molecules compete for the hydrogen bond interactions which hold the two strands of DNA together. By reducing the amount of free water, the Thermotogales reduce the rate of meltdown. Note that this is free water. The amount of total water in the cell is not much reduced. Rather, the ionic strength of the cytoplasm is increased, so that water in the cell is bound up in hydration shells around these ions. (2) Most mesophiles use DNA gyrases, enzymes which help uncoil DNA when it needs to be open for RNA synthesis. Hyperthermophiles have the opposite problem, and have evolved reverse gyrases that effectively tie the DNA up in knots (actually supercoils, not literal knots) so that it can't melt. These enzymes are characteristic of the Archaea. Thermotogales actually have both gyrases and reverse gyrases. (3) Eukaryotic cells use special, arginine-rich proteins called histones and protamines to fold up DNA compactly when it is not in use. Hyperthermophiles use analogous proteins simply to keep the DNA together.
Some important RNA species are also partially double-stranded, for example transfer RNAs and the structural RNAs that are incorporated into ribosomes. These also denature at high temperature. Hyperthermophiles eliminate all mismatches, bulges, and other irregularities that, in mesophiles, lead to flexibility in the RNA. Hyperthermophile RNAs are also usually as short as possible, with no extra sequences, since shorter sequences have fewer nonfunctional folding possibilities. Additional base modifications and changes in protein binding may also be used to stabilize double-stranded regions.
Proteins also denature, but making them resistant to temperature appears to be a relatively simple. However, enzymes are only active over a particular temperature range. Adaptation to high temperature necessarily means that the enzyme will only work at high temperature.
Perhaps the most challenging problem of high temperature is not the effect on macromolecules, but the instability of small, high-energy molecules. As we have mentioned previously, ATP is basic to the energy budget of the cell. Other nucleotide triphosphates (GTP, UTP, CTP, etc) are essential to nucleic acid synthesis, as well as specialized enzyme reactions. But these molecules are quite unstable at high temperatures, and it is simply unknown how hyperthermophiles are able to deal with this challenge.
Image: Hell by Hans Memling (1485) from It's The End Of The World As We Know It...Again
") The Thermotogales are often cited as one of the most basal clades of the Eubacteria. Indeed, even using the most sophisticated techniques, involving statistical analysis of completely sequenced genomes, Thermotogales consistently scores close to the base of the Eubacteria, not far from its presumed origin in the Archaea. Wolf et al. (2001).
The Thermotogales are often cited as one of the most basal clades of the Eubacteria. Indeed, even using the most sophisticated techniques, involving statistical analysis of completely sequenced genomes, Thermotogales consistently scores close to the base of the Eubacteria, not far from its presumed origin in the Archaea. Wolf et al. (2001).
The problem is that we can't be sure that those Archaean genes are "native" to the Thermotogales. Thermotoga is one of the relatively few organisms in which we have really good evidence for large amounts of lateral gene transfer. That is, some genes in Thermotoga have not simply been passed on from parent to daughter cell, they have somehow been acquired from other species. In fact, it has been estimated that 25% of the Thermotoga genome has been co-opted, rather than inherited, from the Archaea. Nesbo et al. (2001). Similar studies have revealed gene transfer and extensive genomic diversity across different strains of Thermotoga. The authors of the current crop of phylogenetic studies express confidence that their results are sufficiently robust not to be misled by lateral gene transfer. Yet, it is difficult to see where this confidence comes from.
 If we may be permitted a speculative leap here, the Thermotogales may be close to the boundary between Darwinian, organismal evolution and an entirely different model of genetic transmission. This model, admittedly, has disturbing resonances in the thoroughly discredited ideas of Trofim Denisovich Lysenko 1898–1976), notably bad scientist and champion of the inheritance of acquired characteristics. [1]
If we may be permitted a speculative leap here, the Thermotogales may be close to the boundary between Darwinian, organismal evolution and an entirely different model of genetic transmission. This model, admittedly, has disturbing resonances in the thoroughly discredited ideas of Trofim Denisovich Lysenko 1898–1976), notably bad scientist and champion of the inheritance of acquired characteristics. [1]
Fortunately, the resonance is misleading. What we are suggesting is not that ordinary evolutionary processes are inapplicable. Rather, we suggest that they are applicable at a different level. If genes can be transmitted separately from genomes, then the appropriate unit of evolutionary change is the gene and not the genome.
Taken to its logical extreme, one may imagine a stromatolite, bacterial mat or biofilm of the deep Proterozoic, in which cells are essentially empty husks, while various genes migrate between them, sorting themselves out into functional communities by natural selection. If a cell happens to contain a complement of genes which work well together, those genes will flourish, multiply, and those genes will be transmitted and enter into other combinations, just as if each gene were an independent species in an evolving ecosystem.
Things could never have been quite that simple; but we are constructing a paradigm here, not a complete picture. Yet recent studies of bacterial ecology in hyperthermophiles suggest that we're not that far off, either. Hot springs and geothermal vents are places of violent activity. Since enzymes and membranes are "tuned" to work best in a particular temperature range, a prudent hyperthermophile will find a location which is frequently at the right temperature and lock itself to some substrate. Thus bacteria in these environments are normally found in tightly bound bacterial mats. What's interesting is that these mats experience both horizontal and vertical gradients of important environmental variables. Horizontally, the average temperature changes quickly with distance from the heat source. Vertically, the availability of light and circulating nutrients varies enormously with depth in the bacterial mat. All of these variables are subject to both cyclic (daily or seasonal) and stochastic (random) fluctuations.
Conventional organismal ecology would lead us to make three predictions in this scenario: (1) the horizontal gradients would lead to species sorting, with successive populations of more thermophilic bacteria being found in a concentric pattern as we approach the heat source; (2) the vertical gradients would lead either to species sorting or to cyclic cell migrations during daily or seasonal cycles; and (3) populations would be relatively similar in similar environments near different springs.
 The third prediction is sometimes true. The first two are untrue. See, e.g., Miller & Castenholtz (2000); Ramsing et al. (2000). What seems to happen instead is that one, or perhaps two, species come to occupy the entire gradient. Yet genetic variability within that species exceeds, by a considerable degree, what would normally be expected in a limited population. For example, Miller & Castenholtz identified four different types of 16S (large subunit) ribosomal RNA along a thermal gradient in a hot spring population of Synechococcus. Ribosomal RNA, as we have mentioned, is one of the most conservative biomolecules in all of life. Because it is extensively folded, and must maintain very precise interactions with numerous ribosomal proteins, viable mutations are exceedingly rare. The possibility that a single population (of anything, much less a bacterium) has independently evolved 4 different alleles of this RNA species seems remote. Even if statistically possible (and it might be), consider how much more likely it is that some other species, already adapted to this temperature zone, would colonize the region and out-compete the evolving Synechococcus. We are almost forced to the conclusion that, in this environment, "Synechococcus" is at most a rather vague phenotypic context and that several rRNA "species", probably acquired from immigrants, are undergoing species sorting independent of this context.
The third prediction is sometimes true. The first two are untrue. See, e.g., Miller & Castenholtz (2000); Ramsing et al. (2000). What seems to happen instead is that one, or perhaps two, species come to occupy the entire gradient. Yet genetic variability within that species exceeds, by a considerable degree, what would normally be expected in a limited population. For example, Miller & Castenholtz identified four different types of 16S (large subunit) ribosomal RNA along a thermal gradient in a hot spring population of Synechococcus. Ribosomal RNA, as we have mentioned, is one of the most conservative biomolecules in all of life. Because it is extensively folded, and must maintain very precise interactions with numerous ribosomal proteins, viable mutations are exceedingly rare. The possibility that a single population (of anything, much less a bacterium) has independently evolved 4 different alleles of this RNA species seems remote. Even if statistically possible (and it might be), consider how much more likely it is that some other species, already adapted to this temperature zone, would colonize the region and out-compete the evolving Synechococcus. We are almost forced to the conclusion that, in this environment, "Synechococcus" is at most a rather vague phenotypic context and that several rRNA "species", probably acquired from immigrants, are undergoing species sorting independent of this context.
And, speaking of sorting, we have wandered a long way from our brief introduction to an obscure Eubacterial clade. The implications of this mode of life are something we will have to sort out elsewhere—perhaps when we reach into the depths of the Archaea. (ATW030330)
[1] On this site, we go to some lengths to avoid anything related to politics. Accordingly, we won't talk about Lysenko's politics. We note simply that his story is an interesting one and that, on a personal note, it is one of the very reasons we do not permit politics on this site. (ATW050729)
Miller, SR, & RW Castenholz (2000), Evolution of thermotolerance in hot spring cyanobacteria of the genus Synechococcus. Appl. Environ. Microbiol. 66: 4222–4229
Nesbo, CL, S L'Haridon, KO Stetter & WF Doolittle (2001), Phylogenetic analyses of two "Archaeal" genes in Thermotoga maritima reveal multiple transfers between Archaea and Bacteria. Mol. Bio. Evol. 18: 362–375.
Ramsing, NB, MJ Ferris & DM Ward (2000), Highly ordered vertical structure of Synechococcus populations within the one-millimeter-thick photic zone of a hot spring cyanobacterial Mat. Appl. Environ. Microbiol. 66: 1038–1049
van Ooteghem, SA, SK Beer, & PC Yue 2001), Hydrogen production by the thermophilic bacterium, Thermotoga neapolitana. Proc. 2001 DOE Hydrogen Prog. Rev.
Wolf YI, IB Rogozin, NV Grishin, RL Tatusov & EV Koonin (2001), Genome trees constructed using five different approaches suggest new major bacterial clades. BMC Evol Biol. 1: 8.
