| Palaeos: | Bones: Dermal Bones | |
| Vertebrates | Palatal Series: Palatines |
| Page Back | Unit Back | Unit Home | Unit References | Unit Dendrograms ("Cladograms") |
Glossary | Taxon Index |
| Page Next | Unit Next | Vertebrates Home | Vertebrate References | Vertebrate Dendrograms | Bones | Time |
|
Bones
|
Dermal Bones
Facial Series |
Palatal Series
|
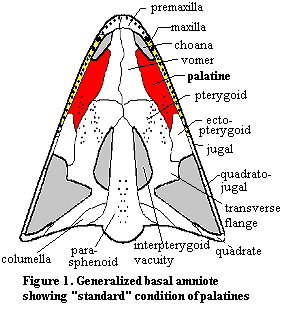 Anatomy: The palatines are dermal bones in the mid-palate. They are derived from neural
crest ectoderm, like many other dermal bones. As has become customary, we will
begin our tour with a visit to Bob, the Basal Amniote. Figure 1.
This time, Bob is shown in palatal view, i.e., looking at the roof of Bob's
mouth with the entire lower jaw removed. Bob, as usual, has the Standard
Condition. In this case, the Standard Condition means that the palatine contacts
the maxilla broadly, along almost its entire posterior length. This is a very
important landmark, since the palatines almost never lose contact with
the maxilla. On the anterior section, the palatines lose contact with the
maxilla and form the edge of the internal nasal openings, the choanae. Medially,
the palatines contact the vomers and the pterygoid. Posteriorly, they terminate
at the ectopterygoid which, despite the name, seem to have developed as one of
several dermal bones on the edges of the palate. That is, the ectopterygoids may
be serially homologous with the palatines.
Anatomy: The palatines are dermal bones in the mid-palate. They are derived from neural
crest ectoderm, like many other dermal bones. As has become customary, we will
begin our tour with a visit to Bob, the Basal Amniote. Figure 1.
This time, Bob is shown in palatal view, i.e., looking at the roof of Bob's
mouth with the entire lower jaw removed. Bob, as usual, has the Standard
Condition. In this case, the Standard Condition means that the palatine contacts
the maxilla broadly, along almost its entire posterior length. This is a very
important landmark, since the palatines almost never lose contact with
the maxilla. On the anterior section, the palatines lose contact with the
maxilla and form the edge of the internal nasal openings, the choanae. Medially,
the palatines contact the vomers and the pterygoid. Posteriorly, they terminate
at the ectopterygoid which, despite the name, seem to have developed as one of
several dermal bones on the edges of the palate. That is, the ectopterygoids may
be serially homologous with the palatines.
Many of the variations on the Standard Condition in more derived tetrapods involve moving the bone more posteriorly. In this case, the ectopterygoids tend to be reduced and the posteromedial edge of the palatine comes to rest on the transverse flange of the pterygoid. The posterior movement of the palatines may be associated with elongation of the anterior jaw. In this case, the maxilla itself develops its own medial flange anterior to the choana and the area between the posterior edge of the choanae and the transverse flange is compressed.
Another frequent variation, particularly in basal forms occurs when the palatine develops a tooth-bearing marginal ridge in apparent serial homology with the maxilla. In some sarcopterygians, for example, the palatines actually bear the larger marginal teeth and the maxilla's tooth-bearing role is "marginalized." On the other side, several different lineages have developed secondary bony palates in which the palatines overgrow the central bones and meet medially.
Functions: As mentioned above, the palatines have an extraordinarily stable association with the posterior maxilla. This surely has some important functional correlate, but it is not easy to see what this might be. It does seem that the palatines are associated with the following special functions.
(a) The lateral roof of the palate: the first and most obvious function is that the palatines for part of the roof of the palate.
(b) Secondary palate: as mentioned above, several very different lineages, including crocodiles and mammals have gone further and adapted the palatines as an important element of a full secondary palate, separating the business of breathing from the business of eating.
(c) Margin of the choana. In choanates, the palatines are strongly associated with the border of the choana. The exact relationship is variable. As we will see, the interaction in, for example, mammals can become quite complex.
(d) Tooth bearing: There are some clear advantages to having big teeth on an interior bone, since the tooth-bearing structure can be supported on both sides. In fact. it is harder to guess why this function was generally abandoned than why it was adopted.
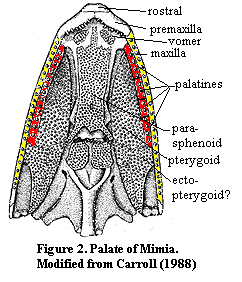 Phylogeny:
Unlike many of the obscure bones in this section, there is an enormous amount of
easily accessible data on the phylogenetic vagaries of the palatines. Obviously,
no vertebrate has palatines until dermal bones appear on the palate. Presently,
the earliest reasonably well-known fish palate is from Mimia, an early
actinopterygian from the Late Devonian Gogo of Australia. The arrangement in Mimia
is weird, but recognizable. See Figure 2. Mimia has
no less than 8 palatines, 4 serial homologues on each side. They are all
marginal, tooth-bearing bones with no detectable roofing function. The lateral
roofing bone is, instead, the pterygoid. The center of the palate is dominate by
a broad, tooth-bearing braincase element, the parasphenoid.
Phylogeny:
Unlike many of the obscure bones in this section, there is an enormous amount of
easily accessible data on the phylogenetic vagaries of the palatines. Obviously,
no vertebrate has palatines until dermal bones appear on the palate. Presently,
the earliest reasonably well-known fish palate is from Mimia, an early
actinopterygian from the Late Devonian Gogo of Australia. The arrangement in Mimia
is weird, but recognizable. See Figure 2. Mimia has
no less than 8 palatines, 4 serial homologues on each side. They are all
marginal, tooth-bearing bones with no detectable roofing function. The lateral
roofing bone is, instead, the pterygoid. The center of the palate is dominate by
a broad, tooth-bearing braincase element, the parasphenoid.
A plate-like dermal bone forming the rim of the adductor chamber in the posterior corners is identified as the ectopterygoid. However, in view of later developments, it may be that the ectopterygoid is actually derived from one of the posterior palatines. In modern actinopterygian fish, as in tetrapods, there is only one palatine per side, and it bears no teeth. The ectopterygoid is immediately posterior and is likewise toothless. However, in sarcopterygians such as Eusthenopteron, a fish close to the tetrapod line, both bones bear marginal teeth. Thus, the palatine of tetrapods and actinopterygians is probably the same bone. However, the tetrapod ectopterygoid is likely derived from one of the ancestral palatine serial homologues. Whether this is homologous to the ectopterygoid of modern fish is unclear.
By the level of Eusthenopteron and Ichthyostega, the palatines already approach the Standard Condition. Both the palatines and the ectopterygoids retain their raised marginal dentition. However, they both have developed substantial roofing processes into the palate. The parasphenoid has retreated, and the pterygoid has become the main roofing bone. The marginal palatine ridge curves inward at the choanae, similar to the Standard Condition except for the large tooth-bearing ridge. The same condition, with minor variations, applies to all of the basal tetrapods, including the temnospondyls. However, in temnospondyls, the interpterygoid vacuities are sometimes so large that the pterygoids are edged out, and the palatines border directly on the vacuities. See, for example the figure at Thoosuchinae.
Lissamphibia are, as usual, so strange that it is not easy to determine what has happened. Oddly, Caecilians make the most sense. As the figure at Eocaecilia shows, this Jurassic form has essentially the Standard Condition, apparently as with other microsaurs. In modern caecilians, the maxilla has developed its own roofing process and the palatines are pushed inward, but still retain large teeth (not merely denticles). In salamanders, the bone is simply lost -- possibly fused with the vomers as is asserted to be the case in Karaurus. In frogs, the palatine is retained, but only as a slight rim around the huge vacuity on each side of the braincase bones of the central palate.
 Back
on the main line of tetrapod evolution, we have reached the level of Bob. Most
forms between anthracosaurs and the Sauria have the Standard Condition, except
that there is a persistent tendency to reduce or fuse the ectopterygoid with the
palatines. As mentioned above, this is associated with compression of the middle
palate, with the palatines pressed back on the transverse process of the
pterygoid. Turtles,
whether considered as parareptiles or aberrant diapsids, show this condition to
a marked degree. Figure 3 shows an example. Note that the
maxillae have developed their own roofing processes, typical of more advanced
tetrapods, and that the palatines are approaching the midline of the palate --
typical of chewing or diving animals that need a strong seal to separate the
breathing and eating functions. All basal diapsids show either the Standard
Condition or one of the same variations exhibited by turtles.
Back
on the main line of tetrapod evolution, we have reached the level of Bob. Most
forms between anthracosaurs and the Sauria have the Standard Condition, except
that there is a persistent tendency to reduce or fuse the ectopterygoid with the
palatines. As mentioned above, this is associated with compression of the middle
palate, with the palatines pressed back on the transverse process of the
pterygoid. Turtles,
whether considered as parareptiles or aberrant diapsids, show this condition to
a marked degree. Figure 3 shows an example. Note that the
maxillae have developed their own roofing processes, typical of more advanced
tetrapods, and that the palatines are approaching the midline of the palate --
typical of chewing or diving animals that need a strong seal to separate the
breathing and eating functions. All basal diapsids show either the Standard
Condition or one of the same variations exhibited by turtles.
Most Sauropterygia and other lepidosauromorphs are likewise unremarkable. One interesting study are the placodonts. The placodont palate bears a rather striking resemblance to the turtle palate except that both the palatines and the maxillae bear huge crushing teeth. However, given the limited range of variation seen in this entire phylogenetic neighborhood, the resemblance could easily be a matter of functional similarity leading to convergent results. The absence of the basisphenoid from the palate of Placodus is also a probably significant difference.
 Snakes,
too are a notable exception to the rule. In fact anomalepidid
and some boid snakes are
the only vertebrates in which the palatines lose contact with the maxilla. Anomochilus
(Figure 4, modified from Cundall
& Rossman (1993)) shows a transitional form. Note how, even in this
highly derived design, the basic relationships of palatine, vomer, pterygoid and
maxilla are preserved, as is the basic architecture of the choana, between the
vomers and the choanal process of the palatines.
Snakes,
too are a notable exception to the rule. In fact anomalepidid
and some boid snakes are
the only vertebrates in which the palatines lose contact with the maxilla. Anomochilus
(Figure 4, modified from Cundall
& Rossman (1993)) shows a transitional form. Note how, even in this
highly derived design, the basic relationships of palatine, vomer, pterygoid and
maxilla are preserved, as is the basic architecture of the choana, between the
vomers and the choanal process of the palatines.
In archosauromorphs, the pattern is a bit different from the Standard Condition in that the palatines are almost always in contact with the transverse flange of the pterygoid. Consequently, the ectopterygoids are pushed out of the way and become fairly marginal elements. At the same time, many crocodylomorphs developed secondary palates which, as mentioned above, tends to involve expanding the palatines medially to meet at the midline. Since the palatines tended to move posteriorly and medially, the pterygoids could not reach the anterior palate. Consequently, crocs tend to elaborate the roofing processes of the premaxilla, anterior maxilla and the vomers.
With the dinosaurs,
things begin to get decidedly more strange. In ornithischians,
the medial migration of the tooth rows completes the separation of the palatines
from the anterior palate  elements.
Figure 5 (modified from Weishampel
& Horner, 1990)) shows the palate of the hadrosaur,
Brachylophosaurus. Here, the pterygoid has been pushed back so far
posteriorly that a portion actually appears as an element of the occipital wall
at the back of the skull.
elements.
Figure 5 (modified from Weishampel
& Horner, 1990)) shows the palate of the hadrosaur,
Brachylophosaurus. Here, the pterygoid has been pushed back so far
posteriorly that a portion actually appears as an element of the occipital wall
at the back of the skull.
Theropods, too have moved the pterygoid to the back of the palate and the vomer is recruited as the central bone of the anterior and middle palate. However, the transformation is not as drastic as in the plant eaters. Even the pattern in the Mesozoic bird, Ichthyornis is still recognizable, with effort. Modern, neornithine birds (Figure 6), however, are difficult to make out based on the Standard Condition. In fact, the avian palate looks quite a bit like an ornithischian palate except that (a) most of the pieces are fused together and (b) the elements are stretched out along the upper jaw to an even greater degree.
 Basal synapsids are but
one step removed from Bob, and thus show the Standard Condition. In fact, the
anterior palate of early proto-mammals looks a bit like a turtle or a
pareiasaur. However there is no maxillary roofing process and the palatines take
up the slack in, for example, Ennatosaurus. This resemblance is increased in the therapsids which do develop
roofing processes of the maxilla.
Basal synapsids are but
one step removed from Bob, and thus show the Standard Condition. In fact, the
anterior palate of early proto-mammals looks a bit like a turtle or a
pareiasaur. However there is no maxillary roofing process and the palatines take
up the slack in, for example, Ennatosaurus. This resemblance is increased in the therapsids which do develop
roofing processes of the maxilla.
The critical change occurs at the level of Thrinaxodon. The palate of Thrinaxodon can be seen "live" at Thrinaxodon CAT Scan Rendering or in a more conventional diagram at Estes, R., 1961 (figure 2). What is happening here is that the palatine has developed a process which is anterior to the choana. In basal cynodonts, this amounts to no more than a little ridge of bone running along the labial edge of the choana. By the level of Thrinaxodon, this ridge has developed its own roofing process which has grown back over the choana as a sort of reflected process of the palatine. In fact, it begins to grow back over the palatine itself. Figure 7 shows the general configuration in a slightly more derived form in the eucynodont, Probainognathus. This species is well on its way to developing a full secondary palate. Both the maxillary and premaxillary roofing processes and the reflected process of the palatines are continuous and meet their opposite numbers at the midline. The choana is completely covered by the reflected palatine at least as far as the level of the orbit.
 This is the basic mammalian pattern. The therian
mammals merely extend this pattern. The roofing processes of the maxillae
grow posteriorly and medially along the full length of the maxilla. The "reflected" palatines are then reduced to smaller, medial elements
around the level of the mid-orbit. This simplicity is deceiving, since the
palatine then develops a complex dorsal process which contributes to the definition of the nasal cavity.
The original palatine is lost and the pterygoid is positioned more posteriorly.
In some forms, notably primates, the remaining palatine elements fuse with the
maxillae to form a continuous bony secondary palate.
This is the basic mammalian pattern. The therian
mammals merely extend this pattern. The roofing processes of the maxillae
grow posteriorly and medially along the full length of the maxilla. The "reflected" palatines are then reduced to smaller, medial elements
around the level of the mid-orbit. This simplicity is deceiving, since the
palatine then develops a complex dorsal process which contributes to the definition of the nasal cavity.
The original palatine is lost and the pterygoid is positioned more posteriorly.
In some forms, notably primates, the remaining palatine elements fuse with the
maxillae to form a continuous bony secondary palate.
Discussion: The palatine is an interesting contrast to the premaxilla. The premaxilla has a number of quite different and exacting functional roles which froze its general form and position quite early in tetrapod evolution. The palatine, by contrast, has far fewer constraints, particularly in those lineages in which it no longer provides a tooth-bearing ridge. Nevertheless it maintains strong contact with the maxilla in virtually all vertebrates. This consistency is hard to understand in functional terms, although it makes reasonably good sense from an embryological perspective.
Another interesting feature is the tendency of both the maxilla and premaxilla to develop their own roofing structures which "compete" with the palatines. This suggests that the maxilla has always had this capability and that the original series of palatines were derived from an earlier generation of maxillary roofing processes. To take the chain of speculation one step further, this suggests that the maxilla itself was originally a flat dermal plate of some kind, possibly homologous to the superognathal of placoderms. ATW 010417.
Links: muscle actions; II. Osteology. 5b. 5. The Palatine Bone. Gray, Henry. 1918. Anatomy of the Human Body.; Dept of Anth: Palatine Page; Skull of the Sea Turtle; PPT Slide.
| Page Back | Unit Home | Glossary | Page Top | Page Next |
checked ATW050819