| Palaeos | Anapsida | |
| Vertebrates | Procolophonidae |
| Page Back | Unit Home | Unit Dendrogram | Unit References | Taxon Index | Page Next |
| Unit Back | Vertebrates Home | Vertebrate Dendrograms | Vertebrate References | Glossary | Unit Next |
|
Abreviated Dendrogram
REPTILIOMORPHA |--SYNAPSIDA `--+--EUREPTILIA | ANAPSIDA |--+--Millerettidae | `--Eunotosaurus |--Mesosauridae `--+--Lanthanosuchoidea `--+--Bolosauridae `--Procolophonia |--+--Nycteroleteridae | `--Pareiasauridae `--Procolophonoidea |--Owenettidae | `?--TESTUDINES `--Procolophonidae |--Sauropareion `--+--Phaanthosaurus `--+--Theledectinae `--+--Tichvinskia `--"Horned procolophonids" |--Procolophoninae | |--Eumetabolodon | `--Procolophon `--Leptopleuroninae |
Contents
Overview |
The Procolophonidae are a group of early herbivorous reptiles of the anapsid lineage that attained a world-wide distribution during the Triassic period. They were small stocky creatures, with blunt chisel-like teeth and triangular skulls. They seemed to come from nowhere following the P-T extinction event, to dominate the Triassic microvertebrates niche.
In fact, the procolophonids had three things going for them: the ability to feed on tough fibrous vegetation, an armour of defensive spines, and the ability to dick and burrow, which, as with modern burrowers, would have surved both to hide from predators and to survive the excessive heat of the day
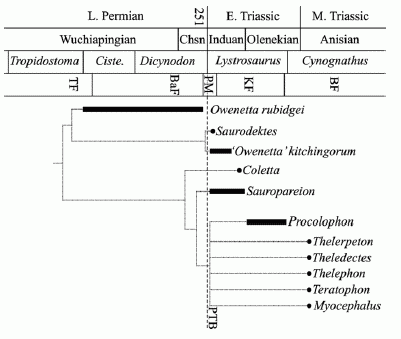 Cladogram of procolophonoids plotted against the South African Permo-Triassic biostratigraphy. Vertical bars and solid circles indicate taxon ranges and single specimen occurrences, respectively. Abbreviations: Chsn, Changhsingian; Ciste., Cistecephalus Assemblage Zone; PTB, Permo-Triassic boundary. Numbers indicate millions of years ago. Diagram and caption from Botha et al 2007 |
Although there are a number of fragmentary Permian remains attributed to procolophonids, the family only is known with certainty from the earliest Triassic. Along with temnospondyls, cynodonts, and the dicynodont Lystrosaurus, earliest Triassic owenettids and procolophonids were important components of the post-extinction recovery fauna. In South Africa three species of procolophonoids (the owenettids Owenetta kitchingorum and Saurodektes rogersorum and the procolophonid Sauropareion anoplus) are known only from the Palingkloof Member of the Balfour Formation, the very lowest part of the Lystrosaurus zone, no more than 30 m above the Permo-Triassic boundary (Botha et al 2007). We are looking at a sudden and amazing evolutionary radiation occuring within only tens of thousand sof years of the main extinct event, perhaps less.
But these sorts of logarithmic differences in timescales are hard to represent diagramatically. This is the reason why the stylised chrono-cladogram, whilst providing very useful information regarding the stratigraphic range of the various taxa, it does not do justice to the suddeness of the procolophonid story. Because logarithmic distinctions in timescales are hard to present against a uniform geological timescale (which deals with increments millions of years or more), the diagram shows what look like (if it is taken literally), a number of ghost lineages reaching back into the Permian. But it is more reasonable to assume that the great procolophonian adaptive radiation really only occurred after the P-T extinction event, in the earliest Triassic. The same thing happened with earliest Cenozoic mammals, as the fossil record unambiguously shows.
It may even be that the evolutionary radiation of survivors into suddenly totally empty niches may have been so rapid as to have taken mere centuries or millennia, or at most tens of thousands of years immediately after the P-T event (or between several extinction pulses). At this scale, where direct ancestors-descendent reklationships are involved, there is no need to avoid paraphyly the way standard cladistics does, because every instance of speciation is by its very nature a paraphyletic process. We know that species can give rise to other species by several means, perhaps the most common being budding (when geographically isolated populations undergo rapid genetic drift); less common would be anagenesis (where one species slowly evolves into a another) and splitting (like anagenesis but with several descendent species). Reading the diagram from a gradist perspective therefore, we would speculate that at the PT-boundary, still in the midst of the extinction event, there is an immediate evolutionary radiation, where Owenetta rubidgei quickly evolves into both Saurodektes rogersorum and Owenetta kitchingorum (which implies that S. rogersorum should perhaps be renamed Owenetta rogersorum). Procolophonids may also have appeared as part of this same adaptive radiation, beginning with a number of early types, before the rise of the very successful, indeed ubiquitous and cosmopoliton Procolophon trigoniceps. A quantum leap beyond its predecessors, larger, better armoured, able to digest tougher plant material, P. trigoniceps dominated Gondwana for all of the middle and late Early Triassic, before giving rise to further species which, although here oversplit in a bewildering array of genera, should probably just beconsiderd further species of the genus Procolophon. In the northern part of Pangea meanwhile, a rival development emerged, the Leptopleuronines. Between them, these two groups would be the rabbits of the Triassic, abundant small burrowing herbivores, pre-adapted to harsh conditions, able to flourish wherever they go MAK120402
The procolophonid's first line of defence was a protective burrow. The hypothesis that procolophonids were burrowers is a popular and persuasive one. It was first suggested with the discovery of Procolophon trigoniceps remains found within a burrow-cast (Groenewald 1991). Since then, many other suggetsions and observations have been made. Leptopleuron lacertinum possesses several morphological traits that are associated with a burrowing mode of life (Säilä 2010a). The related Hypsognathus fenneri lacks cranial kinesis, which may reflect a burrowing lifestyle (Sues et al. 2000). All three known specimens of Koiloskiosaurus coburgensis are preserved together, dorsal up and in the same orientation, which makes sense if the three individuals were occupying a burrow (Botha-Brink and Modesto (2007)). The wide flat body of Procolophon is proportionally quite similar to that of the extant horned lizard Phrynosoma which burrows in loose sand (deBraga 2003). But these are all advanced species. What is most intersting is that from the very start, their most primitive and earliest forms, procolophonids were burrowers. this is evident from Sauropareion anoplus, which we have already seen is one of those little animals that appeared suddenly in the imediarte aftermath of the P-T exutinction event. It exhibits skeletal modifications such as the presence of a spade-shaped skull, robust phalanges, and large unguals, all associated with digging and burrowing (MacDougall et al 2012). While neither large nor armoured, it could certainly conceal itself from preditors.
 Life reconstruction of Procolophon trigoniceps from Cisneros 2006 p.58. In this illustration the quadratojugal processes (cheek bones) are covered by long keratinous spines, and there are also large digging-claws. Keratinous spines are common in large or stocky-bodied modern lizards like Iguana, Phrynosoma, and Uromastyx, and P. trigoniceps and other advancxed procolophonids probably were similarly equipped. Long quadratojugal and supratemporal spines on Procolophon and on leptopleuronines doubtless served as anti-predatory mechanism, as with extant phrynosomatid lizards (Young et al., 2004, cited in Cisneros 2006). |
 Two recent equivalents of the procolophonidae: (1) the Thorny Devil Moloch horridus (Squamata - Agamidae), central and western Australia. Length upto 20cm. These reptiles live almost exclusively on ants Photograph by Wouter!, aka Retuow, via Wikipedia, GNU Free Documentation/Creative Commons Attribution Share Alike |
 (2) the American Desert Horned Lizard or "horny toad" Phrynosoma platyrhinos - (Squamata - Iguania - Phrynosomatidae) of western North America. Length about 10 cm. An ant-eater like Moloch (by means of convergent evolution, as the two are only distantly related) Photo by Chris Brown USGS, (via wikimedia), Public domain |
If you look at a life reconstruction of Procolophon you wonder how such a soft plump looking little animal, easy pickings for a hungry Thrinaxodon, could have survived and flourished worldwide for several millions of years (and the family overall for 50 million). In fact, procolophon probably looked much more like modern spiny desert lizards like the Australian Thorny Devil and American Horned Lizard, shown above.Spines would have provided protection from preditors, and in addition, there was the ability to burrow. Procolophonids had a low broad body, very much like like Phrynosoma but several times as long. In addition, unlike modern thorny or hormed lizards, but like the iguana, they were herbivores; the larger size was probably required for food processed
More efficient food processing was certainly one the keys to procolophonid success. This is revealed in the evolution of progressively more efficient teeth, which doubtless accompanied many soft-body physiological adaptions. While early forms (e.g. Tichvinskia vjatkensis) were still omnivores that would have fed on hard skinned insects as well as tough plant matter, the development of labiolingually expanded crowns enabled a diet of high-fiber plant matter, a niche thatr was convergently exploited by several different evolutionary lines. In the case of the bigger and more advanced forms such as most procolophonines and leptopleuronines, large molar-like teeth developed for processing tough, fibrous foods (Cisneros 2006 p.101) MAK120326
Procolophonidae: Lydekker 1890 (or Cope, 1889)
Phylogenetic definition: All taxa more closely related to Procolophon trigoniceps Owen 1876 than to Owenetta rubidgei Broom 1939. (Cisneros 2006 p.92)
Range: Late Perm? Triassic, Cosm.
Phylogeny: Procolophonoidea : Owenettidae + * : Sauropareion + (Phaanthosaurus + (Theledectinae + (Tichvinskia + (Procolophoninae + Leptopleuroninae)))).
Comments: Here based on cladistic systematics. Of the three traditional subfamilies, Procolophoninae and Leptopleuroninae are monophyletic, Spondylolestinae is paraphyletic (Cisneros 2006 p.108)

Sauropareion anoplus Modesto, Sues and Damiani 2001
Range: Palingkloof Member of the Balfour Formation and lower strata of the Katberg Formation, Beaufort Group, Karoo Supergroup, Induan portion of the Lystrosaurus Assemblage Zone, just above the P-T Boundary (Earliest Induan) of Sth Afr
Phylogeny: Procolophonidae : (Phaanthosaurus + (Theledectinae + (Tichvinskia + (Procolophoninae + Leptopleuroninae)))) + *.
Size: skull length ~3 cm.
Characters: deep posteromedian emargination of the skull table, deep occipital shelves of the parietals and supratemporals, posteroventral facial process of the quadratojugal. (Modesto et al 2001)
Comments: Known from three partial skeletons from Vangfontein, Middelburg District, South Africa. The proportions of the body are similar to those of extant digging animals, and the animal seems to well represent burrowing abilities that would have characterised the procolophonidae as a whole. These include a spade-shaped skull (suitable for moving and packing soil, as with various extant mammals, such as the golden mole, marsupial mole, and mole-rat). While the limbs are not heavily modified for digging, possessing none of the modifications that would increase the mechanical advantage of the limb, such as elongated acromion and olecranon processes found in moles, ground squirrels and aardvarks, which provide a larger area for muscle attachment. But the unguals are relatively large, making them suitable for digging. The non-terminal phalanges are short and robust, which makes the fore and hind feet rigid so they can endure mechanical forces when digging; the modern gopher tortoise has a similar adaptation. While not having the extreme limb modifications seen in some mammals, this would not prevent burrowing abilities (MacDougall et al 2012). Apart from or along with the related Coletta seca (known froma single skull found somewhat higher in the Katberg Formation (Lystrosaurus zone) (Botha &Smith 2006, Botha et al 2007)) this is the most basal procolophonid (Cisneros 2006, Cisneros & Ruta 2010, MacDougall et al 2012). These animals, like the contemporary Owenetta kitchingorum, show that owenettids happily survived the P-T extinction event (Modesto et al 2001).
Graphics Skeletal reconstruction from MacDougall et al 2012. Scale bar equals 2 cm
Phaanthosaurus ignatjevi Tchudinov & Vjuschkov, 1956
Range: Vokhmian Gorizont, Induan, Russia
Phylogeny:Procolophonidae : Sauropareion + ((Theledectinae + (Tichvinskia + (Procolophoninae + Leptopleuroninae))) + *).
Theledectinae Cisneros 2006 : Theledectes perforatus, Eumetabolodon dongshengensis and the Arcadia procolophonid.
Range: Early Triassic of China and Aus,
Phylogeny:Procolophonidae : Sauropareion + (Phaanthosaurus + ((Tichvinskia + (Procolophoninae + Leptopleuroninae)) + *)).
Comments: Cisneros (2006)'s analysis recovered a previously unrecognized clade of procolophonids from south and east Pangaea. This includes a new unnamed procolophonid from the Arcadia Formation of Australia. The Theledectines are small procolophonids distinguished by enlarged, monocuspid marginal teeth with circular bases. These are different from those of Nyctiphruretus or the owenettids, which although they also have circular bases are thinner and sharper (Cisneros 2006 pp.93-4)
Tichvinskia vjatkensis Tchudinov & Vjuschkov, 1956
Range: Rybiskian Gorizont, Olenekian, Russia
Phylogeny:Procolophonidae : Sauropareion + (Phaanthosaurus + (Theledectinae + ((Procolophoninae + Leptopleuroninae) + *))).
Size: length: 25 cm,
Link: Palaeocritti
Unnamed clade ("Horned Procolophonids") : "crown group" [1] procolophonids
Phylogenetic definition: Procolophoninae and Leptopleuroninae (node based) (Cisneros 2006)
Range: Triassic, Cosm.
Phylogeny: Procolophonidae : Sauropareion + (Phaanthosaurus + (Theledectinae + (Tichvinskia + * : Procolophoninae + Leptopleuroninae))).
Comments: This unnamed clade might perhaps be called "Ceratoprocolophonia", as its members can be readily identified by the presence of quadratojugal horns. Also characterized by a more robust build, notably a wide rib cage, and a considerably larger size in comparison to earlier procolophonids and owenettids. (Cisneros 2006 pp.94-5). This clade (including Tichvinskia) showed a significant shifts in diversification rates relative to other anapsids (parareptiles) (Tsuji & Müller 2009), indicating the success of the procolophonid adaptive radiation during the Triassic. MAK120329
Notes: [1] using the cladistic term "crown group" here in a non-anthropocentric and non-neontological context
Procolophoninae Procolophoninae Lydekker 1890 Procolophoninae Seeley, 1888 emend. Romer, 1956, Ivakhnenko, 1979 : Procolophon, Eumetabolodon, Thelerpeton, Teratophon
Phylogenetic definition: All taxa more related to Procolophon trigoniceps Owen 1876 than to Leptopleuron lacertinum Owen 1851. (Cisneros 2006 p.92)
Range: Eartly to Mid Triassic, Sth Afr, China, Ant, Sth Am.
Phylogeny: Procolophonidae : Sauropareion + (Phaanthosaurus + (Theledectinae + (Tichvinskia + ("Horned procolophonids" : Leptopleuroninae + * : Eumetabolodon + Procolophon)))).
Characters: Snout deep and short, postfrontal contacts frontal and parietal only. (Cisneros 2006)
Comments: By inclusion of Procolophon this group represents "typical" procolophonids of the early Triassic. The clade recovered by Cisneros 2006 is not well supported, requiring only one step to collapse. Some of the taxa, such as Kapes, have elsewhere been assigned to the Leptopleuroninae (Säilä, 2008). MacDougall et al 2012 recovered four members of this clade: Procolophon trigoniceps, Thelerpeton (Thelegnathus) oppressus, Teratophon (Thelegnathus) spinigenis (these three of the Early-Middle Triassic Cynognathus zone, (Olenekian-Anisian) of South Africa), and Eumetabolodon bathycephalus. This is also the same as a one-step suboptimal scenario by Cisneros 2006 ("clade L").
Eumetabolodon bathycephalus Li, 1983
Horizon: Upper He Shang Gou Formation (Early Triassic), Shaanxi and Nei Monggol, Inner Mongolia, North China.
Phylogeny: Procolophoninae : Procolophon + *).
Size: skull length: 6 cm
Characters: short high skull, short snout, forward positioned pineal foramen ("third eye") partially bordered by frontals, and long, posterior (rearward) located jaw articulation are all unique features among procolophonids (Lucas 2001 p.99).
Comments: Known from numerous skulls and skull fragments, Eumetabolodon bathycephalus is a typical member of the Procolophonid family, similar to the Gondwanan Procolophon and the European Koiloskiosaurus, all charactrerised by classic features such as triangular skull, large orbits, and a small number of transversely broad, bicuspid teeth. The 18 known skulls of Eumetabolodon fall into four size classes and allow tooth replacement during ontogeny to be analyzed. The study by Li (1983) shows evidence of tooth replacement during growth, in which conical teeth postcanine teteh in juveniles (implying an insectivorous diet) were replaced by transversely broad, bicuspid teeth in adults (implying a herbivorous lifestyle); hence the name "changeable tooth". (Lucas 2001 p.99). Two species have been placed here; E. bathycephalus is the best known taxon, whilst another species, E. dongshengensis has also been described based on a poorly preserved partial skull. Cladistic analysis by (Cisneros 2006 indicates that Eumetabolodon is a paraphyletic or polyphyletic genus: Eumetabolodon bathycephalus is a procolophonine, whereas E. dongshengensis is a theledectine and must be placed in a new genus (Cisneros 2006 p.108)
Link: Palaeocritti
Procolophon: Owen 1876
Synonyms: Procolophon baini Broom 1905, Procolophon brasiliensis Cisneros and Schultz 2002, Procolophon cuneiceps Seeley 1878, Procolophon griersoni Seeley 1878, Procolophon laticeps Seeley 1878, Procolophon minor Owen 1876, Procolophon platyrhinus Seeley 1905, Procolophon pricei Lavina 1983, Procolophon sphenorhinus Seeley 1905 (ref Paleobiology Database)
Range: Early Triassic (Middle Lystrosaurus zone, Induan to Early Cynognathus zone, Olenkian), South Africa, Antarctica, and Brazil (pan-gondwanan).
Horizons. Lower Katberg Formation (upper Lystrosaurus zone) to lower part of the Burgersdorp Formation ((Cynognathus Subzone A = lowermost Langbergia Subzone, Late Induan-Olenekian). of the Karoo Basin, Fremouw Formation of the Transantarctic Basin (Kitching et al. 1972; Colbert and Kitching 1975) and the Sanga do Cabral Formation of the Parana Basin in Brazil (Barberena et al. 1981; Lavina 1983). (Cisneros 2006 pp.35, 47-8)
Phylogeny: Procolophoninae : Eumetabolodon + *.
Size: Length: 30 cm
Comments: The eponymous representative of the Procolophonidae (to say nothing of the Procolophonoidea, the Procolophonia, and the Procolopnonomorpha...), known from hundreds of specimens, most of them found in the Lower Triassic sediments of the South African Karoo, where it is outnumbered only by the ubiquitous Lystrosaurus, this long-lived and widespread species can quite plausibly be assumed to have evolved from smaller, more primitive, stratigraphically earlier Triassic forms like Sauropareion, which it would have replaced or supplanted.
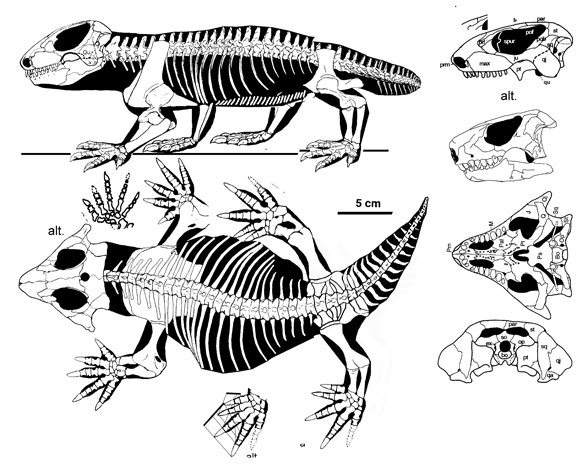
In the past included under paraphyletic orders like the Cotylosauria (Watson 1914, Colbert, 1969, Kuhn, 1969) and Captorhinida Carroll, 1988 it is now considered a representative of the anapsid or parareptile lineage. While too specialised to be directly ancestral to turtles, it still may be close to the turtle line in the interpretation we have followed here (Reisz and Laurin 1991), although the question of turtle origins remains controversial and it may be that the procolophonoidea alone (Lee 1995) or the parareptilia as a whole Rieppel & deBraga 1996 are an entirely extinct lineage
Like other advanced procolophonids, Procolophon possesses a robust, wide torso (imagine an oversized desert horned "toad") and enlarged chisel-like teeth, suggesting a high-fibre diet (Gow, 1977, Hotton et al. 1997). Limb adaptations show Procolophon was capable of burrowing (deBraga 2003), and this species is most likely the author of burrows whose solidified casts have been recovered from the Lystrosaurus Assemblage Zone (Groenewald 1991).
The prominent horn-like quadratojugal processes on the side of the skull give the head a distinctive triangular shape. Unusually but not uniquely for early anpasids, some individuals have temporal openings, which have in the past been interpreted as different species (Hamley & Thulborn 1993).
As fossils are known across a wide geographical range, from the South African Karoo to the Transantarctic Mountains to the Parana Basin in Brazil, it can reasonably be assumed that the actual distribution of the living animal was across the whole of in Gondwana. P. trigoniceps persists into the latest Early Triassic (Early Cynognathus Zone), co-existing with the diapsid Palacrodon and the temnospondyl amphibian Trematosuchus. Although more than half a dozen species have been recognised in the past, including recently P. pricei and P. brasiliensis from the contemporary (Lystrosaurus assemblage age) Sanga do Cabral Formation of Brazil (Cisneros & Schultz 2002), these are now understood as synonyms or variants of a single species, P. trigoniceps
Later in the Cynognathus zone (early Anisian, earliest middle Triassic), Procolophon's place is taken by a confusion of poorly described and repetitatively named taxa such as Theledectes, Thelegnathus, Thelerpeton, Thelephon, with a single Teratophon to add a little nomenclatural diversity to the mix. Cladistic analysis (Cisneros 2006) resolves Thelerpeton, and Teratophon as close to Procolophon, so it is not unlikely that would represent variations of a single chronospecies. Theledectes appears in tehs ame analysis as an unrelated, more primitive form. Thelephon seems to be part of a transitional clade related to or a basal member of the the leptopleuronines, and Thelegnathus is a polyphyletic taxon or a nomen dubium.
References: Cisneros 2006 pp.35, 47-8, 95-6,
Links: Palaeocritti, Wikipedia, Paleobiology Database MAK120327
Graphics Skeletal reconstruction © David Peters, used with permission
| Page Back | Unit Home | Page Top | Page Next |
page MAK120326
{kind=link}