| Palaeos | Anapsida | |
| Vertebrates | Pareiasauridae |
| Page Back | Unit Home | Unit Dendrogram | Unit References | Taxon Index | Page Next |
| Unit Back | Vertebrates Home | Vertebrate Dendrograms | Vertebrate References | Glossary | Unit Next |
Editor's note: much of the following has been copied as is from kheper net and needs to be integrated more with the Paleos com format. There may also be some overlap in the following text, some of which has been incorporated and doubtless improved by ATW. A small amount of new material has been added. MAK120323

The Pareiasaurs were large late Permian herbivores. They were heavy ungainly animals, frequently the size of an ox, with capacious bodies, strong limbs, broad feet, and thick solid skulls ornamented with bizarre knobs and ridges. They were armoured with bony scutes or osteoderms set in the skin, The largest types were 3 metres long and about 1 1/2 meters tall, with weights exceeding a tonne.
 The
leaf-shaped multi-cusped teeth (left) resembled those of iguanas, caesids,
and other reptilian herbivores. This dentition, together with the
deep capacious body (which obviously housed an extensive digestive tract)
are proof that these fearsome-looking animals were in reality inoffensive
herbivores.
The
leaf-shaped multi-cusped teeth (left) resembled those of iguanas, caesids,
and other reptilian herbivores. This dentition, together with the
deep capacious body (which obviously housed an extensive digestive tract)
are proof that these fearsome-looking animals were in reality inoffensive
herbivores.
Pareiasaurs appear very suddenly in the fossil record. It is clear that these animals evolved from Nycteroleterids, perhaps a Rhipaeosaur-like form to fill the large herbivore niche (or guild) that had been occupied early in the Permian period by the Caesid pelycosaurs and before then the Diadectid amphibians and Edaphosaur reptiles. They are much larger than the diadectids, more similar to the giant caseid pelycosaur Cotylorhynchus (Colbert 1965 p.52). In fact it may well have been the extinction of the Caesids created an ecological vacuum that enabled the Pareiasaurs to appear and suddenly diversify as rapidly as they did (within the span of only two million years). Although the last Pareiasaurs were no larger than the first types (indeed, many of the last ones became smaller), there was a definite tendency towards increase of armour as the group developed.
It has been often suggested that these animals were semi-aquatic. Skeletons have been found in a standing position, indicating that the animals became mired in the mud and swamps while feeding on vegetation. Their remains are also consistently found in channel stream deposits (riverbeds). Of course, this does no mean that they were semi-aquatic; and actually the fact that the animals became stuck argues against aquatic adaptations (a semi-aquatic animal would not likely become stuck, whereas a terrestrial animal would; e.g. the remains of sauropod dinosaurs, which were as terrestrial as elephants, are sometimes found in this position). A more plausible explanation is that these were fully terrestrial animals that frequented watercourses in order to get at the succulent aquatic vegetation.
It has been argued that Pareiasaurs may have evolved into turtles. They had turtle-like skull features, and several genera had bony plates in the skin, possibly the first signs of a turtle shell. However, the case for turtle ancestry is not proven.
Recent reports of a slender stapes (a sound-conducting middle ear ossicle) indicate that pareiasaurs had an efficient ear drum (tympanum) and were able to hear high frequency airborne sounds. MAK000124
References: Benton 2000, 2005, Carroll, 1988 p.205; Colbert19651965 pp.52-3; Fenton & Fenton 1958 p.306; Kuhn, 1969, Laurin, 1996 , Zittel & Eastman 1932 , pp.241-243
Remains of Pareiasaurs are quite common in the Lower Beaufort Series of South Africa, and a great many generic and specific names were originally proposed. It would be very unlikely that such a large number of similar species would co-exist in the same time and space, and it is now recognised that many of these different types are actually the result of individual variation, growth stages, geological deformation of fossils, etc. Now no more than five genera are recognised from South Africa, four from Russia, two from Europe and one from China. MAK000124 Because of the influence of current cladistic nomenclature there is a tendency now toawards monotypal genera (i.e. allowing each genus only a single species); we have resisted this in favour of consistency with non-paleoherpetological and non-vertebrate paleontological taxonomy MAK120329
Pareiasauridae Lydekker 1889 = Pareiasauridae Cope, 1896 (= Pareiasauria, Hallucicrania) : Herbivorous anapsids, some very large (up to 3m).
Range: Middle to Late Permian of Africa, Europe, Russia, Sth Am, & China.
Phylogeny: Procolophonia : Nyctiphruretus + (Procolophonoidea + (Nycteroleteridae + * : Bradysaurus + Velosauria.))
 The beautifully preserved complete remains of the intermediate-grade Russian pareiasaur Deltavjatia vjatkensis. In life this animal would have had a posture very similar to the closely related South African genus Bradysaurus (see mounted skeleton, below). And as with the Bradysaurus, a large number of complete skeletons of these animals have been found in association; the exact causes of for these mass-mortality asemblages is still not clear. Image © Albert Khlyupin and the Vyatka Palaeontological Museum, used with permission. |
Characters: Short, laterally expanded skull, short, foliate teeth; probably herbivorous; palate strongly integrated with braincase; $ palate raised above level of tooth row; transverse process of pterygoid bears marginal teeth; $ the transverse process is at 45 deg. to the midline (rather than 90) and curved ventrally; interpterygoid vacuities closed; choanae not parallel with maxilla; jaw articulation well anterior to occipital condyle; $ lacrimal excluded from narial margin (unclear if this is correct, much less synapomorphic); $ frontal excluded from orbit & in contact with post-frontal; tendency to form flat dorsal surface to head, with orbits on sides & some fusion of dorsal dermal bones; post-parietals fused; supratemporal with large boss; strange, "melted" rugose dorsal cranial surface with projections downward from lower jaw and laterally projecting quadratojugal; $ < 21 presacral vertebrae; caudal vertebrae not posteriorly bent; gastralia absent; deep body with strong ribs; extremely long scapula with long, blade-like acromion process; stocky limbs, with epipodials relatively vertical and facing forward; metapodials wider than long(!); pelvis mammal-like, with ilium dorsoventrally elongated, and short, retroverted pubis & ischium; dorsal buttress over acetabulum; extensive body osteoderms (dermal scutes) in some species.
Further Systematic Description: The skull is short, broad (laterally expanded), massive, and strongly sculptured and knobby. The sutures are not clearly distinguishable. The quadratojugals (cheekbones) project laterally downwards (almost like the hanging "jowls" on a cartoon bulldog-type character). The orbits (eye sockets) are relatively small, and laterally (side of the head) placed. There is an opening for the pineal body ("third eye", use to regulate metabolism in the case of seasonal extremes). the brain-case is long and low, the ear-cavity separated by bone from the brain cavity. The tabular and post-parietal bones of the skull are on the dorsal (upper) surface only. The jaw articulation is well forward of the occipital condyle (the connection between the skull and vertebra), increasing the mechanical efficiency of the jaw musculature while decreasing the gape.
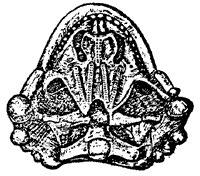 Pareiasaurus palate from Kuhn 1969 |
The palate (left) strongly integrated with the base of the braincase and the skull margins. The bones of the palate are fused, and bear several series of small teeth. The teeth are conical with leaf-shaped crowns. Those on the margins of the jaws area arranged in a close regular series.
There are about eighteen presacral (neck and trunk) vertebrae, each with deep articulation for the ribs, and generally four sacral vertebrae. The sacral vertebrae are unfused (an inefficient arrangement). There are thirty caudal (tail) vertebrae with the anterior ones bearing short ribs. Generally the tail is quite short.
The pelvic bones are massive and ossified. The scapula is much longer than in other primitive tetrapods, and the pelvis has an almost mammalian configuration. This may have helped accommodate muscles that moved the rear limb in a manner similar to the fore and aft gait of mammals.
The humerus is short, massive, and expanded at the ends; the ulna very stout. The hind limbs are slightly shorter than the fore-limbs. The stocky limbs are held close to the body to support the weight. Both fore and hind feet have five digits, which are equipped with broad claws. MAK000124
 |
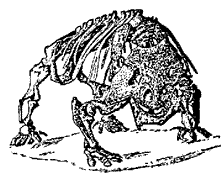 |
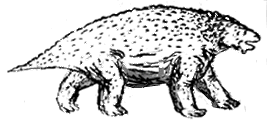 The advanced pareiasaur Scutosaurus karpinskii, above left, Carroll, 1988, p.205; original, 2 meters long; right, front view Kuhn, 1969, p.69; left, sketch on the basis of skeleton above left, by M.Alan Kazlev 1998 The advanced pareiasaur Scutosaurus karpinskii, above left, Carroll, 1988, p.205; original, 2 meters long; right, front view Kuhn, 1969, p.69; left, sketch on the basis of skeleton above left, by M.Alan Kazlev 1998 |
|
Discussion: It is easy to lose track of the main points in the wealth of detail about pareiasaurs poured out in recent years. Whether or not Permian pareiasaurs gave rise to turtles, were a sister group of turtles, or would sooner shoot their sister than be related to a turtle, they were not turtles. Neither were they simply aberrant lizards or Paleozoic proto-bovines or terrestrial placodonts. They were, from all appearances, rather unique products of the late Paleozoic radiation of amniotes. In common with other lost branches of early radiations, they show a mosaic of characteristics and show some similarity to all of the above.
Let us, for a moment, forget both Cenozoic models and cladistics and look at them for what they were as animals. They had a short, blunt jaw. The skull was generally boxy, particularly by Paleozoic standards. Likely this was a function of jaw specializations. These herbivores needed big mouths, since vegetation is relatively nutrient poor. They had also developed a short jaw and deep skull in order both lengthen the jaw muscles and to get a more vertical angle on the lower jaw with bigger muscle mass. The pareiasaurs also developed some dental specializations: nothing fancy, just flattened, foliate teeth along the jaw while retaining palatal teeth. The pterygoid had to assume a new and more complex shape as a result of the other changes. The palate as a whole became more tightly integrated with the skull, and this consolidation and buttressing is even reflected in a tendency to fuse some of the dermal skull elements, particularly near the midline and at the "new" breakline where the top of the skull curved down to form the sides.
In life, the skull might not have reflected these changes on casual inspection because the posterior dorsal elements are flared out and ornamented with lumps, horns, bosses, and a remarkable variety of other motifs for protection, display, solar panels or something completely unknown. It also seems rational to suppose that, like many herbivores, they possessed cheeks and, given the large flare, potentially enormous cheeks. As a result, the possible functions of the laterally expanded skull might relate to volume as well as just dorsal area. By way of speculative example, one might consider: fat storage, an elaborate salivary digestive system, mechanisms for hearing or sound production, or the oral equivalent of a gizzard. By way of analogy, pelecaniforms also use large gular pouches for feeding young and in cooling behaviors, both entirely plausible behaviors for pareiasaurs. None of these are, of course, supported by a shred of evidence. They are listed solely to suggest that conventional models based on living creatures may be inadequate.
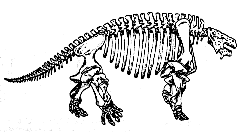
 The primitive pareiasaur Bradysaurus baini - reconstructed skeleton, from the Museum of Natural History in Berlin (Museum für Naturkunde) Photo by Masur, Wikipedia, GNU Free Documentation/Creative Commons Attribution Share Alike |
The limb structure of pareiasaurs is also a peculiar mosaic. The limb girdles have been elongated. Undoubtedly one reason for this was simply to allow the enormous gut to clear the ground. However, most extant animals solve this problem by lengthening the limbs themselves. Pareiasurs had elongated limb girdles, but relatively short, squat limbs. Much has been made of the mammal-like shape of the pelvis, but the comparison may be over-played. An organism with this general shape cannot move effectively using only lateral undulations like a lizard or a salamander. It follows that the limbs, however locomotion is accomplished, have to translate their force along the central axis. Thus, selection will necessarily favor a rigid body and limb girdles that act, as much as possible, by exerting force on the vertebral column. Hence, the scapula is very long and think, the ribs are stout, the vertebral number is reduced, the pelvis is slanted forward and the iliac crest lies parallel with the vertebral column, the tail (no longer much connected with locomotion) is short and wide. The limbs themselves are still perhaps closer to Ichthyostega than to mammals or dinosaurs. The epipodialia are not as short as some illustrations make out, but the metapodia certainly are. Perhaps the length of the girdle elements was designed precisely to prevent and dampen torsion while making the most of the relatively small vertical motions possible with a lizard-like leg structure. By contrast, the feet are surprising. The astragalus has overgrown the calcaneum to such an extent that there is essentially only one tarsal, and the digits have a strangely dinosaurian look to them. These are not exactly long-distance runners, but not just outdoor furniture, either.
In short, Pareiasaurs make sense, but on their own terms. We might imagine a specimen of medium-large size, under 2m long and perhaps 200kg, as a moderately selective low browser. It moves slowly about in rolling, fairly open country, working steadily away at a huge mouthfuls of leaves, occasionally shaking its head in an odd fashion to redistribute the load and clear away insects. Its head looks far larger than its skull would suggest, inflated by pendulous cheeks. It bears an irregular, patchy pattern of scutes on its broad back and sides, with smaller elements on its limbs. Skin and scutes are of contrasting earth tones, breaking up its outline in the brush and tall ferns.
Had they survived the end-Permian extinction better, they might have competed well with Ornithiscian dinosaurs or even mammals. Perhaps they did survive -- as turtles. Whatever the case, pareiasaurs are best understood as a group unto themselves, rather than an early model of anything that walks today.
Note:This taxon has been one of the battlegrounds of one of the more interesting taxonomic battles of recent years, with the respective forces led by (among others) MSY Lee (turtles are Pareiasaur sisters) and Olivier Rieppel (turtles are part of a third major diapsid lineage) These Notes follow the more orthodox path, with some serious doubts expressed elsewhere. ATW030308
Links: Introduction to Pareiasauria, Pareiasauria: More on Morphology; dinosaurs- scutosaurus karpinski; Phylogeny and Classification of Amniotes; Turtle Origins; South African Museum - Fossil Reptiles of the South African Karoo; PANGEA (Italian); pareiasaur; Provelosaurus; Provelosaurus reconstruction; animals. ATW030308
Some associated clades:
Hallucicrania Lee 1995. The clade Hallucicrania was coined by MSY Lee, for Lanthanosuchidae + (Pareiasauridae + Testudines) (see links: Tree of Life Phylogeny and Classification of Amniotes and Hallucicrania [Pareiasauriformes] after Lee, 1993, 1996 and 1997 ...). Lee's pareiasaur hypothesis is looking rather less likely following the discovery of Odontochelys, a transitional aquatic turtle with very non-pareiasaur-like teeth and whose half shell matches embryonic development in modern testudines (an intriguing case of Haeckel's ontogeny recapitulates phylogeny). From another angle, recent cladistic analyses reveal Lanthanosuchids to have a much more basal position in the Procolophonomorpha , and that the nearest sister taxon to the pareiasaurs are the rather unexceptional and conventional looking Nycteroleterids (Müller & Tsuji 2007, Lyson et al 2010) the two being united in the clade Pareiasauromorpha (Tsuji et al 2012). The weird dermal sculpturing in both pareiasaurs and lanthanosuchids, that inspired Lee to coin such a wonderful name, is therefore the result of convergence. It would be a real shame though to discard such a wonderful name as Hallucicrania. For now at least we're keeping it as a synonym for Pareiasauria. ATW030402 updated and expanded MAK120323.
Pareiasauroidea Nopsca, 1928. The clade Pareiasauroidea (as opposed to the superfamily or suborder Pareiasauroidea) was used by Lee 1995 for Pareiasauridae + Sclerosaurus. More recent cladistic studies place Sclerosaurus in the procolophonid family Leptopleuroninae (Cisneros 2006, Sues & Reisz 2008) which means the similarities with pareiasaurs are the result of convergences
Pareiasauria Seeley, 1988. If neither Lanthanosuchids or Testudines are included in the clade, the Pareiasauria only contains the monophyletic family Pareiasauridae. There seems little reason to use Seeley's ordinal-style name. For this reason we have reverted to the traditional linnaean term. MAK120323.
References: deBraga & Rieppel (1997); Lee (1997).
Links: Introduction to Pareiasauria; South African Museum - Fossil Reptiles of the South African Karoo; final run.PM6; E V (Dutch -- excellent materials); cordados répteis primitivos (Spanish); Turtle Origins; PANGEA (Italian). ATW030308. Palaeocritti - Pareiasauria (brief but useful coverage of every species) MAK120401
 Bradysaurs: Watson 1914 (monotypal Subfamily Bradysaurinae Huene 1948)
Bradysaurs: Watson 1914 (monotypal Subfamily Bradysaurinae Huene 1948)
Synonyms: Brachypareia, Bradysuchus, Koalemasaurus, Platyoropha
Stratigraphic range: Lower to Upper Tapinocephalus zone, Lower Beaufort Beds, of the Karoo basin, South Africa; most common in the Lower Tapinocephalus zone (Lower to Middle Tapinocephalus zone in Boonstra, 1969 pp26, 32,; Lower Tapinocephalus to Pristerognathus (= old Upper Tapinocephalus) zone in Ward et al 2005, supplem. info, range data; both = Capitanian age)
Phylogeny: Pareiasauridae : Velosauria + *.
Characters: single row of vertebral scutes
Size length about 2.5 meters long, weight about half a tonne to a tonne (Fenton & Fenton 1958 p.306; Colbert 1965 p.52).
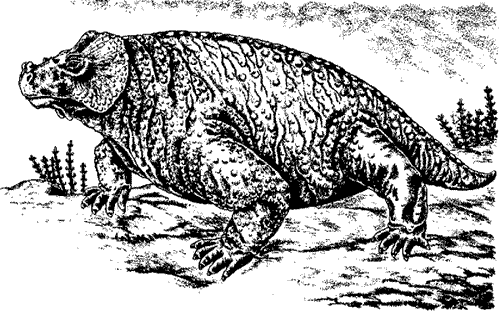
Comments: Despite its large size, Bradysaurus is actually the most primitive known genus of Pareiasaur. The Bradysaurs can be considered the ancestral type from which the others developed, even if it lived at the same time as one or two other genera. It can be assumed that Bradysaurus evolved somewhat earlier in time, during the Wordian epoch (Eodicynodon zone). Fossils are rare in the Eodicynodon zone, which may explain its absence.
The skull is large, broad, flattened, and rounded at the front. The skull-bones are coarsely sculptured and knobby, the sutures between them not clearly visible. The marginal teeth are high-crowned with only a few cusps, this being a primitive characteristic. The cusps consist of a terminal group of three cusps, the anterior with one to three and the posterior two to three cusps.
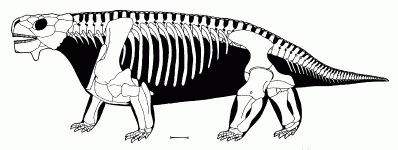
The phalangeal count is 2,3,3,3,2 on the fore-foot and 2,3,3,4,3 on the hind. The whole body is covered in thin dermal scutes, which are quite smooth over the back. The armour is not as heavily developed as in later forms.
The top of the ilium is vaulted. There are 5 cervical, 15 dorsal, 4 sacral, and about 39 tail vertebrae.
Kuhn lists no less than nine species for this genus, but this is certainly an excessive number. Boonstra distinguishes only four species on the basis of tooth structure, two of which Kuhn places in the genus Embrithosaurus. I have decided to take the more cautious position and only have two species in this genus. MAK000124
With the genus Bradysaurus, this clade contains only B. seelyi. Lee (1997) argues that the only available material from other putative species of this genus, B. baini, lacks any distinguishing autapomorphies. However, B. seelyi is closely related to Nochelesaurus and Embrithosaurus and they are treated here as a paraphyletic stem group of all primitive pareiasaurs except Deltavjatia. ATW
Graphics Skeletal reconstruction from Lee 1997, life reconstruction from Fenton & Fenton 1958 p.306
 |
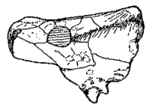
Bradysaurus baini (Seeley 1892)
Synonyms: Pareiasaurus baini Seeley, 1892, Platyoropha broomi Haughton & Boonstra, 1929, Bradysaurus broomi (Haughton & Boonstra, 1929)
Stratigraphic range: Tapinocephalus zone (Capitanian), Lower Beaufort Beds, Karoo basin, South Africa
Comments: This is the type species for the genus. The quadrajugal region (cheek-bones) are only moderately developed. The snout is broad and rounded. There are 15 or 16 pairs of overlapping teeth in each jaw. The genera Brachypareia, Bradysuchus, Koalemasaurus, Platyoropha would also seem to belong here. Platyoropha and Brachypareia are indistinguishable from Bradysaurus, and Brachypareia based on apparently immature specimens (Boonstra, 1969 p.30), although Brachypareia watsoni is now considered a synonym of Embrithosaurus angustus, Bradysuchus, is based on a pathological feature. The skull illustrated is 42 cm long MAK000124
Tsuji & Müller 2009 in their unpublished analysis make this the most basal species of pareiasaur MAK120325
Graphics illustration of skull from Kuhn 1969
 |
Bradysaurus seeleyi Haughton and Boonstra 1929
Synonyms: Bradysaurus vanderbyli Haughton & Boonstra, 1929
Stratigraphic range: Tapinocephalus zone, Lower Beaufort Beds, South Africa
Characters: - Lee 1997
Comments: Boonstra considered this a valid species of Bradysaurus. The cheekbones are greatly enlarged and heavy. There are 19 or 20 pairs of strongly overlapping teeth on each jaw. This species clearly was not as common as B. baini. Length of specimen at left 44 cm MAK000124
According to Lee 1997 this is a "metaspecies" and lacks unique characteristics that can be used in phylogenetic analysis. Tsuji & Müller 2009 in their unpublished analysis place it between B. baini and as the most basal species of pareiasaur MAK120325
Links: Introduction to Pareiasauria; The first Karoo Reptiles and their origin; Victoria west; Paleontology and Geology Glossary- Br; Bradysaurus (German); Bradysaurus (skull photos); museum für naturkunde berlin- bradysaurus baini; UCMP Mystery Fossil Number 13; South African Museum - Fossil Reptiles of the South African Karoo; South African Museum - Trace Fossils of the Ancient Karoo; ALBANY MUSEUM - KAROO FOSSILS (view the Tapinocephalus Zone landscape); Bradysaurus. ATW031009.
Graphics illustration of skull from Kuhn 1969
| Page Back | Unit Home | Page Top | Page Next |
checked ATW040118, revised MAK120325. All text by ATW placed in the public domain. All text by MAK Creative Commons Attribution license. This does not cover illustrations, which are copyright their original authors or publishers. Older material by MAK originally appeared on Kheper net