| Palaeos: | The Proterozoic Eon | |
| Proterozoic | Proterozoic 1 |
| Page Back | Back: Archean | Up: Precambrian | Unit Home |
| Page Next | Next: Phanerozoic | Down: Paleoproterozoic | Timescale |
The Proterozoic is, roughly speaking, the time when more or less "modern" plate tectonics began to govern over other processes in determining the form of the Earth's crust. Although continents were small, they consisted of stable cratons. Mid-ocean spreading ridges did a good deal of the moving, just as they do today. However, everything happened a good deal faster. The magma on which the continents floated was hotter, less viscous, and closer to the surface. Hot spots were probably hotter. The continents moved more swiftly, collided more often and tended to fracture or suture with greater frequency.
Life developed from the infant stage of single celled organisms to an adolescence of Eukarya and early plants, fungi, animals. Perhaps other forms developed as well which we know less about because they failed to explode in the Cambrian Explosion. Like all other adolescents, Life grew much larger, discovered sex, and changed its mind frequently about what it was going to be when it grew up. Undoubtedly it tried out many forms and lifestyles which, had we learned of them at the time, we would have sternly disapproved. Life engaged in risky behaviors, such as carelessly spewing so much oxygen into the atmosphere that it nearly poisoned itself until it learned to adapt. It moved out from the warm geothermal vents where, perhaps, it was raised, and nearly froze to death once or twice by wandering into very serious Ice Ages without its mittens. Somehow, in spite of a number of these very close calls, it grew up into the sort of grown-up Life we know today.
ATW040930. Public Domain. No rights reserved.
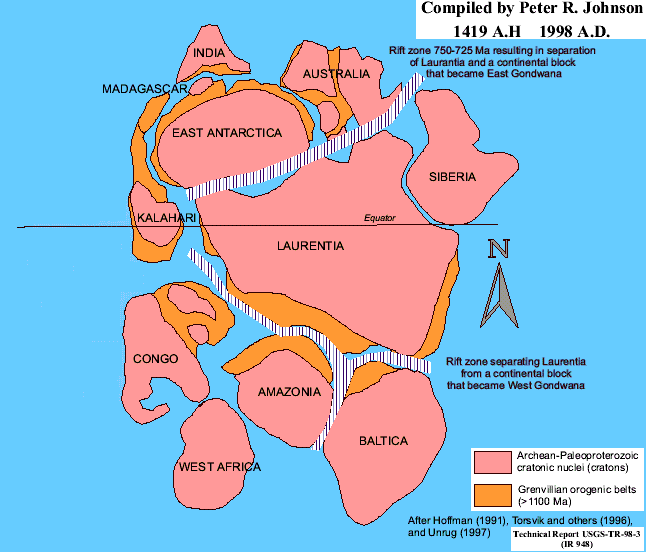
We know very little of the geography of the Proterozoic. As Dr. Christopher Scotese notes on his paleomap site: "With available data, 650 million years is about as far back as we can go." Of course, this has not stopped him (or many others) from pushing the paleomap envelope to about 750 Mya. However, beyond that there are only educated guesses.
One fact now does seem reasonably clear. About 1100 or 1200 Mya, most of the landmass of the Earth was locked up in a continent called Rodinia. How Rodinia assembled is speculative at this point. Since Dr. Scotese is undoubtedly the best educated of the educated guessers, you may be interested to see his concept of Rodinia at its greatest extent (perhaps 1100 Mya).
The map above is from The Geological History of Jamestown, Rhode Island. It represents one popular interpretation of the makeup of Rodinia at perhaps 900 - 1000 Mya. Most of the continents are rough blobs. However, we do know that Laurentia was approximately upside down compared to the present day. Therefore the center of the Earth at that time was somewhere in upstate Minnesota -- which is enough to prove that this was truly a very different world from our own.
About 900 Mya, Rodinia started to fracture; and the pattern
of fragmentation has become fairly well known. This map sets up a very
nice structure around the Amazonia - Baltica - Laurentia corner which is known
as a three-armed graben. This is not some sort of con game or high
tech slot machine. It is a true continent-busting confluence of two faults
of exactly the type which tore Pangea apart 800 My later and which is trying
(but probably failing) to tear up East Africa in the present day. In fact,
the Kalahari - East Antarctica - Laurentia corner may be the site of a second
graben, just as there were really two such structures involved in the demise of
Pangea. Scotese, seems to place most emphasis on the northwest to
southeast rift splitting Rodinia apart. The huge bulk of Laurentia and
Siberia then rotates clockwise about 120° clockwise as it moves down almost to
the South Pole and 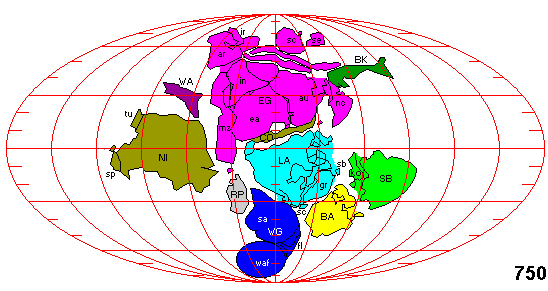 back up again, hitting West Africa from the South just
as East Gondwana (i.e. North Rodinia), having rotated a bit clockwise, hits it
from the Northeast.
back up again, hitting West Africa from the South just
as East Gondwana (i.e. North Rodinia), having rotated a bit clockwise, hits it
from the Northeast.
One could imagine a more peaceful scenario in which Laurentia and Baltica simply cut through the "strait" between Kalahari and East Antarctica, while rotating (either way) to reach their Cambrian positions and orientations. There are excellent reasons why this cannot be the case, not the least of which is the far southern paleolatitude of North America (Laurentia) during the latest Neoproterozoic. Nevertheless, in the spirit of being obnoxious, we offer this additional map, from Steven Dutch of the University of Wisconsin. The large, dark green thing in the west on Dutch's map is the Niger block -- actually pieces of both Africa and South America. See Johnson & Rivers (2004) which supports this this placement. If this version -- not so very different from the map above -- is correct, Laurentia would have a tough time rotating all the way south around these obstacles to reach a Cambrian position roughly where the Niger block is shown, all in only 230 My. Not impossible, mind you, but not really the most parsimonious interpretation from a geometrical point of view.
ATW040930. Text public domain. No rights reserved.
|
Eon |
Era |
when began
My ago |
duration
My |
|
| Proterozoic | Neoproterozoic
1000 mya
|
Ediacaran | 630 | 88 |
| Cryogenian | 850 | 250 | ||
| Tonian | 1000 | 150 | ||
|
Mesoproterozoic
1600 Mya
|
Stenian | 1200 | 200 | |
| Ectasian | 1400 | 200 | ||
| Calymmian | 1600 | 200 | ||
| Paleoproterozoic
2500 Mya
|
Statherian | 1800 | 200 | |
| Orosirian | 2050 | 250 | ||
| Rhyacian | 2300 | 250 | ||
| Siderian | 2500 | 200 |
 The Proterozoic includes over half the history of life on this planet,
but the available data about Proterozoic organisms are exceedingly sparse.
We usually have a little discussion about plankton at about this point on the
Time pages, but we know practically nothing about plankton in the Proterozoic.
Consequently, we'll talk mostly about acritarchs instead. And what exactly is an acritarch? No one is quite sure.
Acritarchs are like sharks' teeth from the Phanerozoic or cigarette butts from the
1960's. That is, they are found everywhere, are practically
indestructible, and come in a variety of interesting shapes and sizes.
They look informative, but ultimately tell us rather little about the organisms
or ecosystem which produced them -- other than the fact that it contained sharks
or smokers, as the case may be. Acritarchs tell us even less because we
don't know what produced them. The typical statement in the literature is: "Acritarchs
[are] a group of decay-resistant organic-walled vesicular microfossils ... .
Most acritarchs from the Proterozoic and Paleozoic are interpreted as unicellular
photosynthetic protists ... ."
Huntley et al.
(2006).
The Proterozoic includes over half the history of life on this planet,
but the available data about Proterozoic organisms are exceedingly sparse.
We usually have a little discussion about plankton at about this point on the
Time pages, but we know practically nothing about plankton in the Proterozoic.
Consequently, we'll talk mostly about acritarchs instead. And what exactly is an acritarch? No one is quite sure.
Acritarchs are like sharks' teeth from the Phanerozoic or cigarette butts from the
1960's. That is, they are found everywhere, are practically
indestructible, and come in a variety of interesting shapes and sizes.
They look informative, but ultimately tell us rather little about the organisms
or ecosystem which produced them -- other than the fact that it contained sharks
or smokers, as the case may be. Acritarchs tell us even less because we
don't know what produced them. The typical statement in the literature is: "Acritarchs
[are] a group of decay-resistant organic-walled vesicular microfossils ... .
Most acritarchs from the Proterozoic and Paleozoic are interpreted as unicellular
photosynthetic protists ... ."
Huntley et al.
(2006).
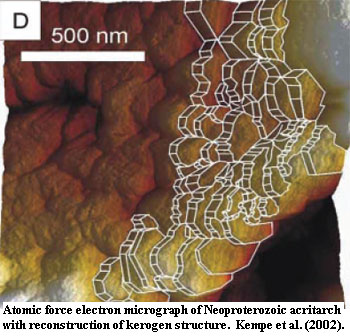
Although acritarchs have been probed and prodded with almost every sort of device known to man, they have yielded relatively little detailed structural information. They are clearly unmineralized, organic-walled structures. After sitting around for one or two billion years, almost all nano-scale molecular organization has been lost. What is left is kerogen, amorphous platelets of polycyclic aromatic hydrocarbons with no obvious resemblance to any familiar cell wall material. Kempe et al. (2002).
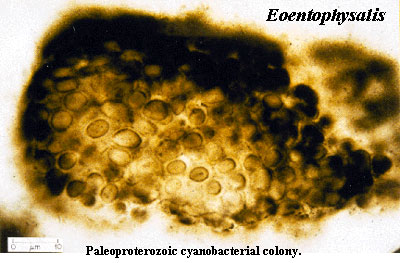
No acritarchs are yet known from the earlier Paleoproterozoic. Biomarker evidence (i.e. the presence of long-chain 2-methylhopanes) suggests that a phytoplankton population was present, consisting mainly/entirely of cyanobacteria. Summons et al. (1999); Falkowski et al. (2004); Canfield (2005).
 Acritarchs
are present in the later Paleoproterozoic; but they are rare and consist of morphologically simple
spheres (i.e. sphaeromorphic acritarchs).
Javaux et al.
(2004). They sometimes look a good deal like
prasinophytes (basal green
algae), and perhaps that's what they are. More likely, they're below the
split between red algae and green algae, like
glaucophytes, or
even basal to all crown eukaryotes.
Many or most acritarchs from the Paleoproterozoic and earliest
Mesoproterozoic
are probably akinetes or other bacterial remains.
Golubkova &
Raevskaya (2005). Akinetes are inactive resting stages of cyanobacteria
induced by cold, lack of food, or similar environmental conditions unfavorable
for growth -- the sorts of stimuli that generally lead organisms to seek
metabolic stasis and/or admission to graduate school.
Meeks et al.
(2002) (review).
Acritarchs
are present in the later Paleoproterozoic; but they are rare and consist of morphologically simple
spheres (i.e. sphaeromorphic acritarchs).
Javaux et al.
(2004). They sometimes look a good deal like
prasinophytes (basal green
algae), and perhaps that's what they are. More likely, they're below the
split between red algae and green algae, like
glaucophytes, or
even basal to all crown eukaryotes.
Many or most acritarchs from the Paleoproterozoic and earliest
Mesoproterozoic
are probably akinetes or other bacterial remains.
Golubkova &
Raevskaya (2005). Akinetes are inactive resting stages of cyanobacteria
induced by cold, lack of food, or similar environmental conditions unfavorable
for growth -- the sorts of stimuli that generally lead organisms to seek
metabolic stasis and/or admission to graduate school.
Meeks et al.
(2002) (review).
Other types of fossils from near the Paleoproterozoic-Mesoproterozoic boundary include coiled or worm-like forms such as Grypania, and possibly related structures resembling beads on a string. Porter (2004). Some of the Grypania-like fossils approach 1 mm in width, which seems unreasonably large for a bacterium or even a colonial bacterial structure. Yet the earliest Grypania are about a billion years too old to be actual worms. So perhaps -- for lack of any other hypothesis -- these are very early Eukarya, possibly outside the crown group.
 By the
early Mesoproterozoic (Calymmian), the
evidence for a eukaryotic grade of organization becomes more definite.
This judgment is based on: "(1) wall structure and surface ornamentation (2)
processes that extend from vesicle walls (3) excystment structures (openings
through which cysts liberate their cellular contents) (4) wall ultrastructure
and (5) wall chemistry."
Javaux et al.
(2004). In particular, large cells with processes extending beyond the
wall (i.e. acanthomorphic acritarchs) are thought to be impossible
without a eukaryotic cytoskeleton. Id. Oddly enough, this
agrees reasonably well with recent "molecular clock" work, which likewise places
the primary radiation of the
Plantae in the Calymmian.
Yoon et al.
(2004). Mesoproterozoic acritarchs include specimens with new
morphological features: ellipsoidal shape, vesicle pores, and a multi-celled or
colonial appearance.
Huntley et al.
(2006).
By the
early Mesoproterozoic (Calymmian), the
evidence for a eukaryotic grade of organization becomes more definite.
This judgment is based on: "(1) wall structure and surface ornamentation (2)
processes that extend from vesicle walls (3) excystment structures (openings
through which cysts liberate their cellular contents) (4) wall ultrastructure
and (5) wall chemistry."
Javaux et al.
(2004). In particular, large cells with processes extending beyond the
wall (i.e. acanthomorphic acritarchs) are thought to be impossible
without a eukaryotic cytoskeleton. Id. Oddly enough, this
agrees reasonably well with recent "molecular clock" work, which likewise places
the primary radiation of the
Plantae in the Calymmian.
Yoon et al.
(2004). Mesoproterozoic acritarchs include specimens with new
morphological features: ellipsoidal shape, vesicle pores, and a multi-celled or
colonial appearance.
Huntley et al.
(2006).
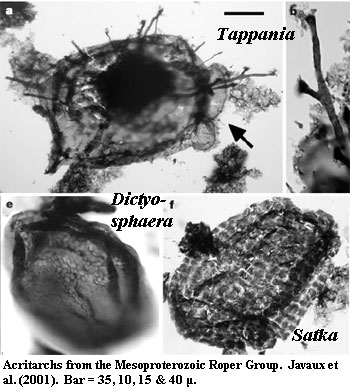
Javaux et al. (2001) studied well-dated samples from the Roper Group of northern Australia. They recovered specimens of the controversial, but almost certainly eukaryotic, Tappania. Note the relatively large size and the long, irregular processes which penetrate the outer wall. A bacterial origin isn't completely impossible, but the more likely explanation is a eukaryote with a well-developed cytoskeleton. In any case, some of the acritarchs known from this era are relatively enormous, such as Chuaria circularis (better known from the Neoproterozoic), which can approach 1 mm in diameter. Golubkova & Raevskaya (2005).
Another key finding of this study was that the Roper acritarchs showed clear ecological zonation, with different populations characteristic of inshore, nearshore, and distal shelf environments. The authors speculate that communities were limited by nutrient mineral runoff, since abundance and diversity seem higher in marginal marine settings. However, there is no guarantee that acritarch diversity reflects biotic diversity generally.
Slightly later (Ectasian) communities from the Ruyang Group of North China are
dominated by Dictyosphaera, an acritarch also found in the
 Roper
Group.
Kaufman & Xiao (2003). This cosmopolitan distribution is typical of
Proterozoic acritarchs. The authors performed ion microprobe isotopic
analysis of individual specimens, and were able to make a rough estimate that
Mesoproterozoic CO2 levels were between 10 and 1000 times higher at
present.
Roper
Group.
Kaufman & Xiao (2003). This cosmopolitan distribution is typical of
Proterozoic acritarchs. The authors performed ion microprobe isotopic
analysis of individual specimens, and were able to make a rough estimate that
Mesoproterozoic CO2 levels were between 10 and 1000 times higher at
present.
It isn't clear that these fossils are crown eukaryotes (i.e. descendants
of the last common ancestor of all living eukaryotes). It seems likely, if
only because all acritarchs are assumed to have been photosynthesizers; but a
good deal of room for doubt remains. However, there is a modest consensus
that crown eukaryotes had appeared by Stenian, if not
earlier, since these latest Mesoproterozoic acritarchs more closely resemble
modern algae.
Javaux et al. (2004);
Porter (2004).
In particular, Bangiomorpha looks like an extant alga and shows a very
unusual "intercalary" pattern of cell division that is characteristic only of
living bangeacean red algae.
Porter (2004). Some acritarchs from this period can be
found with surface ornamentation, and not simply processes. The Stenian
also marked the first
 appearance of stalked cyanobacteria.
Golubkova &
Raevskaya (2005).
appearance of stalked cyanobacteria.
Golubkova &
Raevskaya (2005).
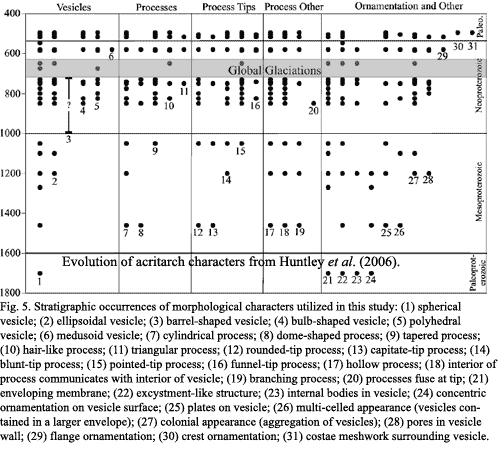
Fossils from even the earliest Neoproterozoic (Tonian) include forms commonly identified as fungi and recognizable modern orders of green algae. Falkowski et al. (2004); Golubkova & Raevskaya (2005). (We omit detailed consideration of metazoans, including Ediacaran forms, and metaphytes, which appeared in the later Cryogenian and Ediacaran. Peterson & Butterfield (2005). These are discussed extensively below, and in the sections devoted to those eras). Morphological features new to the Neoproterozoic include "polyhedral vesicles, bulb-shaped vesicles, barrel-shaped vesicles, triangular and hair-like processes, funnel-tipped processes, processes that fuse at the tips, and flange ornamentation about the vesicle equator." Huntley et al. (2006) (examples and internal citations omitted).
Generally speaking, the data from the Cryogenian are poor, but suggest continuity with the Tonian forms without dramatic changes in morphology or even diversity. Porter (2004). The first fossil remains of testate amoebae appear late in the Cryogenian -- the first good evidence of heterotrophic Eukarya. Porter (2004). The Ediacaran introduced two new communities, one associated with the Ediacaran animals (or whatever they may have been), followed by one associated with the transitional metazoans of the Doushantuo type. The latter include recognizable modern orders of red algae. Xiao et al. (2004). Many acritarchs from this period bore regular processes and surface ornamentation, e.g. Appendisphaera, Ericiasphaera. Golubkova & Raevskaya (2005).
A number of efforts have been made to quantify the pattern of acritarch diversity across the Proterozoic. Knoll (1994); Porter (2004); Huntley et al. (2006). In general they verify the that, after an initial burst of diversification in the later Paleoproterozoic, development was gradual or even static until the Neoproterozoic. Surprisingly, evidence for a diversity or even abundance bottleneck as a result of the Cryogenian "snowball earth" episodes is weak to almost non-existent. See also Olcott et al. (2005) (biomarker evidence). It might be fairer to say that the diversity curve began to take off in the Tonian and was, at most, slowed down in the Cryogenian.
ATW061208. Text public domain. No rights reserved.
 In the long term, the most successful animal group of the Proterozoic were the sponges. We are somewhat reluctant to speak much about Proterozoic sponges because we developed some very peculiar and idiosyncratic ideas about the evolution of sponges in another part of Palaeos. Those speculations have been summarized elsewhere in what is, just possibly, the worst-written essay on this site. We tentatively stand by what was said, if not how we said it.
In the long term, the most successful animal group of the Proterozoic were the sponges. We are somewhat reluctant to speak much about Proterozoic sponges because we developed some very peculiar and idiosyncratic ideas about the evolution of sponges in another part of Palaeos. Those speculations have been summarized elsewhere in what is, just possibly, the worst-written essay on this site. We tentatively stand by what was said, if not how we said it.
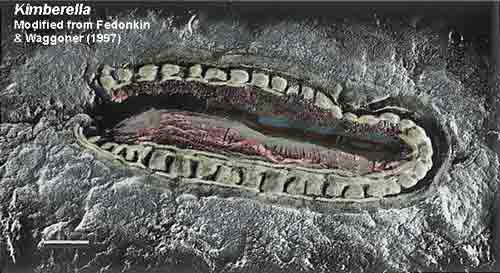
Here we will be more conservative, as we are summarizing the current state of a huge topic about which no two scientists seem to agree. We will introduce only two home-made ideas. The first deals with the phylogenetic position of Kimberella. The second is yet another take on the famous embryos of Doushantuo mentioned in the previous section. The plan (if we can stick with it) is to avoid too much heavy thinking and concentrate on the thoughts of more qualified people.
We will begin with a brief orientation which covers the first 95% of the Proterozoic in a couple of paragraphs. We will then cover the Ediacaran fauna, which we will call "Vendobionta," following the terminology introduced by Seilacher. We will put special emphasis on on the "rangeomorph" vendobionts and some ideas developed by Narbonne (2004). We will briefly apply those ideas to Kimberella, a vendobiont often compared to mollusks. In brief, we suspect that, whether or not Kimberella actually is a basal mollusk, it can also be interpreted as a not-particularly derived rangeomorph vendobiont. After this, we will talk a little about Precambrian sponges as a sort of warm-up to the Doushantuo demilitarized zone. Finally, we will walk gingerly through the Doushantuo minefield itself. The Weng'an locality of the Doushantuo Formation has been the site of ferocious scientific combat since February 6, 1998, when Li et al. (1998) announced the discovery of small sponge body fossils and embryos from Weng'an. We take no sides in this free-for-all, but we introduce a possibility which (so far as we know) none of the combatants seems to have considered: that the embryos of Weng'an are the embryos of vendobionts. ATW081205
 In the old days of molecular phylogeny, roughly speaking the decade between 1995 and 2005, it was common to read "molecular clock" papers confidently announcing that Metazoans had evolved in the Mesoproterozoic, if not earlier. Fortunately, it is no longer necessary for us to spend pages tearing into these contentions. More recent papers, using vastly more sophisticated techniques, arrive at results which are considerably more consonant with the fossil record. Peterson et al. (2008) is a good example, and a good study.
In the old days of molecular phylogeny, roughly speaking the decade between 1995 and 2005, it was common to read "molecular clock" papers confidently announcing that Metazoans had evolved in the Mesoproterozoic, if not earlier. Fortunately, it is no longer necessary for us to spend pages tearing into these contentions. More recent papers, using vastly more sophisticated techniques, arrive at results which are considerably more consonant with the fossil record. Peterson et al. (2008) is a good example, and a good study.
Peterson's group takes considerable care to calibrate their molecular analysis with multiple time points from the fossil record. They find (with generous error bars) that the Fungi diverged from Metazoa at some point in Stenian or Tonian time. We use "Metazoa" to mean toads > toadstools. This is over-inclusive, and we continue to use it only for the sake of consistency with other parts of Palaeos. When most people use the term "animal" they generally mean something more like sponges + Spinoza. In that sense, animals began to evolve in the Cryogenian, with the evolution of the principal sponge clades. Peterson et al. find that most of the high-level animal clades then diverged in the Ediacaran and radiated in the Cambrian. When evaluating these results, it is important to recall that we are speaking of genetic divergence times. Fossils with detectably clade-specific characteristics ("apomorphies") may not appear until long after a clade has diverged genetically from all others. Given that understanding, the results reported by Peterson's group are entirely credible. Since they are closely constrained by the fossil record, such studies represent a synthesis of the currently available molecular and paleontological data.
 Some results from paleontology have also suggested an early start for Metazoa. The more promising of these candidates have been reviewed by Fedonkin (2003). Of these, one of the most interesting is the very simple, beads-on-a-string creature, Horodyskia. Fedonkin & Yochelson (2002). Horodyskia is not uncommon in bottom sediments of Calymmian age (Mesoproterozoic, c. 1500 Mya) or younger in North America and Australia. The conical beads, or "zooids," are plainly connected by something, presumably a stolon. The strings seem to be of relatively uniform length. Zooid size is uniform for each string, but varies considerably between strings, reflecting zooid growth with time. Proportional spacing between zooids is maintained, presumably by resorbtion of alternate zooids. Thus the putative young strings have many small zooids, while older strings are the same length, but have a small number of larger zooids. This kind of coordinated growth suggests, as Fedonkin and Yochelson note, a "tissue-level" organization rather than a colony of individuals.
Some results from paleontology have also suggested an early start for Metazoa. The more promising of these candidates have been reviewed by Fedonkin (2003). Of these, one of the most interesting is the very simple, beads-on-a-string creature, Horodyskia. Fedonkin & Yochelson (2002). Horodyskia is not uncommon in bottom sediments of Calymmian age (Mesoproterozoic, c. 1500 Mya) or younger in North America and Australia. The conical beads, or "zooids," are plainly connected by something, presumably a stolon. The strings seem to be of relatively uniform length. Zooid size is uniform for each string, but varies considerably between strings, reflecting zooid growth with time. Proportional spacing between zooids is maintained, presumably by resorbtion of alternate zooids. Thus the putative young strings have many small zooids, while older strings are the same length, but have a small number of larger zooids. This kind of coordinated growth suggests, as Fedonkin and Yochelson note, a "tissue-level" organization rather than a colony of individuals.
Horodyskia doesn't really enter into our equation. Fedonkin (2003) loosely refers to Horodyskia as a "metazoan." Few seriously assert that Horodyskia is descended from the last common ancestor of frogs and fungi -- much less that it is an "animal" in the sense defined above. Some workers are still not completely convinced that Horodyskia is even eukaryotic. Knoll et al. (2006). Cooperative multicellularity evolved independently in at least six living eukaryote groups (red algae, green algae, 2-3 groups of chromists, slime molds, fungi, metazoans). The surprise is that there are so few known extinct multicellular clades.
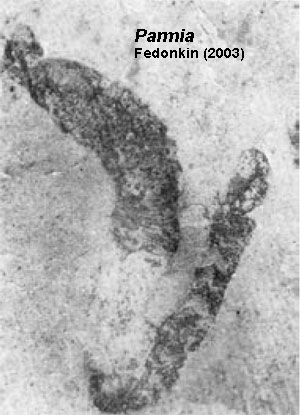
Fedonkin (2003) also mentions the worm-like Parmia and Sinosabellidites, two worm-like organisms of early Neoproterozoic age. Parmia looks like an unholy cross between a sea cucumber (Holothuroidea) and an annelid worm. It is known only from the Tonian of the Russian Platform. Sinosabellidites and several similar forms are slightly younger organisms from China. All show closely spaced annulations, on the order of 200µ apart. However, Dong et al. (2007) interpret organisms of this type as algae, with any apparent proboscis reinterpreted as a holdfast. In short, the evidence for advanced metazoans before the Ediacaran is not compelling. ATW081205.
The environment in which Ediacaran metazoans evolved is only a little less controversial. Some of the issues are discussed above, in the section on acritarchs. The Ediacaran begins with the end of the "Snowball Earth" episode of the Marinoan Glaciation, at about 630 Mya. The current feeling seems to be that this was followed by an increase in atmospheric oxygen or, at least, in oxygenation of the oceans. The main point of contention is whether the oxygenation occurred in some stepwise fashion -- through a series of rapid "events" (Fike et al., 2006; 2007; Scott et al., 2008) -- or proceeded more gradually (Grey & Calver, 2007; Shen et al., 2008).
 The ocean floors where animals developed were quite unlike those of the Phanerozoic. Mobile herbivores were exceedingly rare or absent, as were burrowing worms. Consequently, the sea floor was often covered in an algal/bacterial mat. Much of the mud and bottom ooze of the Phanerozoic is the product of constant churning by animal life. Likewise many of the hard substrates were created by corals and other calcifying animals. Before animals, the usual bottom was a firm, living bacterial blanket, with occasional, usually small, carbonate outcrops built by calcifying bacteria. Dornbos et al. (2005).
The ocean floors where animals developed were quite unlike those of the Phanerozoic. Mobile herbivores were exceedingly rare or absent, as were burrowing worms. Consequently, the sea floor was often covered in an algal/bacterial mat. Much of the mud and bottom ooze of the Phanerozoic is the product of constant churning by animal life. Likewise many of the hard substrates were created by corals and other calcifying animals. Before animals, the usual bottom was a firm, living bacterial blanket, with occasional, usually small, carbonate outcrops built by calcifying bacteria. Dornbos et al. (2005).
The first metazoans lacked any obvious way to move, and also lacked any obvious mouth, gut, or central cavity. Consequently, it is unclear what they ate, or how they fed themselves. McMenamin (1998) argued that the earliest animals hosted symbiotic photosynthetic algae. This hypothesis is currently out of favor because many of the best known animal fossils of the Ediacaran appear to come from deep water -- well below the depth where sunlight could penetrate. Peterson et al. (2003). One currently popular belief is that these organisms absorbed dissolved organic carbon compounds directly from seawater. Sperling et al. (2006). This would require some fairly peculiar ocean chemistry, but many workers find it entirely plausible. ATW081206.
| Page Back | Unit Home | Page Top | Page Next |
