| Palaeos: | Ediacaran Period | |
| Neoproterozoic | Ediacaran - 2 |
| Page Back | Back: Cryogenian | Back: Mesoproterozoic | Up: Neoproterozoic | Unit Home |
| Page Next | Next: Cambrian | Next: Paleozoic | References | Timescale |
555 Ma: Ediacara HillsMaterial from the Ediacara Hills (Flinders Ranges) has still not been precisely dated; it is assumed to be approximately coeval with the White Sea fossils, in the region of 555 Ma (see below), but it could be as young as the +1 to +2 d13C interval, dated at 549 to 543 Ma in southern Namibia. Martin et al. (2000: 844). It is the assemblage from this site that is most widely associated with the base of the Ediacaran biota. Although best known for the 'classical' body fossils, the region also provides interesting traces. One ichnotaxon, similar to that from Zimnie Gory, has been interpreted as the radula scratchings of a mollusc (possibly Kimberella). |
Trace fossil from the Flinders Range |
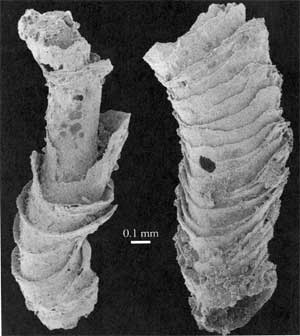
549 to 543 Ma: Nama GroupThe Nama Group is a thick (> 3 km) shallow marine and fluvial foreland basin succession, partitioned into northern and southern sub-basins by an intervening arch, across which most stratigraphic units thin, located in southern Namibia. The age range of the Ediacaran assemblages from the Nama Group is the interval 548.8 ± 1 to 543.3 ± 1 Ma. Grotzinger et al. (1995). In addition to typical Ediacaran taxa, such as the cosmopolitan Pteridinium, the shelly fossil Cloudina first appears slightly below the earliest Ediacaran fossils, extends throughout the Ediacaran range, and into the Cambrian. Moreover, a second, unnamed, shelly taxon ("goblet-shaped shelly fossils") coexists with Cloudina from at least 545 Ma through into the Cambrian. Id. |
Cloudina |
Ediacaran LifeThe following passage is from Benchley and Harper, Palaeoecology, pp.121-123
|
|
| There can be little doubt, on the basis of trace evidence alone, that bilaterian metazoans existed in the Ediacaran, and possibly early in the Ediacaran. Although some traces are simple, rather featureless, winding trails, "others display transverse rugae and contain pellets that can be interpreted as of fecal origin. The bilaterian nature of these traces is not in dispute. Furthermore, such traces must have been made by worms, some of which had lengths measured in centimetres, with through guts, which were capable of displacing sediment during some form of peristaltic locomotion, implying a system of body wall muscles antagonized by a hydrostatic skeleton. Such worms are more complex than flatworms, which cannot create such trails and do not leave fecal strings." Valentine (1995: 90). Sets of paired hypichnial ridges strongly hint at an arthropod s.l. presence. Unfortunately, it is equally true that the relatively few body fossils known from the late Precambrian do not shed much light on the sequence of evolutionary advances that led to the famously diverse Cambrian taxa. There are a few sign-posts, however:
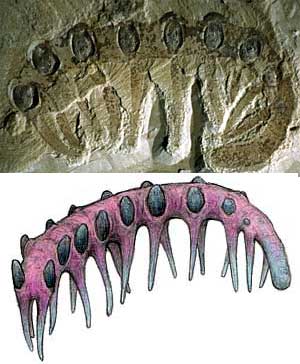
Mineralised skeletons of uncertain affinity - the 'small shelly fauna' - appear just before the beginning of the Cambrian, ~550 Ma, increasing in numbers and diversity towards the Tommotian. The most common skeletal materials are calcium carbonate (aragonite or calcite) and varieties of calcium phosphate. Many of the latter may originally have been carbonates, phosphatized during preservation. The oldest of these to occur abundantly are Cloudina and the allied genera comprising the family Cloudinidae: small, conical fossils made of calcium carbonate, first (?) appearing in the Ediacaran Stirling Quartzite of California (Langille, 1974) and persisting into the Cambrian. Anabarites and Cambrotubulus are other Ediacaran SSF taxa, known from Siberia and Mongolia. While it is not known what kind of organism produced Cloudina, and many other SSFs are equally problematic, some of the Cambrian representatives have been tied back to a firm systematic placement, such as Microdictyon, which is now known to be an onychophoran. |
|
ExtinctionsAdapted from The Precambrian and Vendian Mass Extinctions: Extinctions are proposed to have affected even life's earliest organisms. About 650 million years ago, seventy percent of the dominant Precambrian flora and fauna perished in the first great extinction. This extinction strongly affected stromatolites and acritarchs, and was also the predetermining factor that encouraged the diversification of the Ediacarans. The first extinction of the Precambrian, which largely affected stromatolites and acritarchs, has been correlated with a large glaciation event that occurred about 600 million years ago. This event was of such severity that almost all micro-organisms were completely wiped out. However, this distinct fauna may also have perished in a second extinction event at the close of the Ediacaran. This event may have been responsible for the ensuing diversification of the Cambrian shelly fauna. The Ediacaran extinction, occurring near the close of the Ediacaran period, is currently under debate as to whether an extinction event occurred or not. Many paleontologists believe that the Ediacaran fauna were the progenitors of the Cambrian fauna. However, others believe that the Ediacaran fauna have no living representatives. Under this latter hypothesis, the Ediacaran fauna is believed to have an undergone an extinction, after which the Cambrian fauna evolved. Until more information can be collected, details on the Ediacaran extinction event will remain open to debate.
Although some taxa are now known to have persisted, and others may have evolved into different forms, most of the Ediacarans simply vanish from the fossil record near the beginning of the Cambrian. Some believe this is evidence of a mass extinction. Moreover, in "the past few years, evidence has accumulated for a remarkable perturbation in the carbon cycle close to the Proterozoic-Cambrian boundary. Globally distributed sedimentary successions document a strong (7 to 9 per mil) but short-lived negative excursion in the carbon-isotopic composition of surface seawater at the stratigraphic breakpoint between Ediacaran-rich fossil assemblages and those that document the beginning of true Cambrian diversification. The causes of this event remain uncertain, but the only comparable events in the more recent Earth history coincide with widespread extinction – for example, the Permo-Triassic crisis, when some 90% of marine species disappeared, is marked by an excursion similar to but smaller than the Proterozoic-Cambrian boundary event. An earliest Cambrian increase in bioturbation shuttered the taphonomic window on Ediacaran biology. Thus, while Chengjiang and Sirius Passet fossils indicate that Ediacaran-grade organisms were not ecologically important by the late Early Cambrian, biostratigraphy admits the possibility that Ediacarans were eaten or outcompeted by Cambrian animals. It is biogeochemistry that lends substance to the hypothesis that Ediacaran and Cambrian faunas are separated by mass extinction." Knoll & Carroll (1999). One school of thought holds that Ediacarans may have been largely wiped out by a supposed nutrient crisis – ‘Kotlin Crisis,’ see Brasier (1992) – immediately prior to the Ediacaran-Cambrian boundary. However, other researchers observe that a mass extinction event is not necessary to explain the disappearance of the Ediacarans from the fossil record; conditions may simply have ceased to be favorable to their preservation with the arrival of more numerous and more diverse scavenging and bioturbating organisms. Indeed, the lower boundary of the Cambrian is now defined by the occurrence of a distinctive horizontal burrow trace fossil, Trichophycus (formerly Phycodes) pedum in the reference section at Fortune Head, southeastern Newfoundland. "We cannot tell how abruptly the Ediacaran Faunas became extinct, but only a very small number are represented by possible survivors... ." "Although most Ediacaran fossils have no post-Proterozoic record, they were not immediately succeeded in lowermost Cambrian rocks by diverse crown group bilaterians. Earliest Cambrian assemblages contain few taxa, and the diversity of trace and body fossils grew only over a protracted interval. Hyoliths and halkieriids (extinct forms thought to be related to mollusks), true conchiferan mollusks and, perhaps, chaetognaths enter the record during the first 10 to 12 million years of the Cambrian, but crown-group fossils of most other bilaterian phyla appear later: the earliest body fossils of brachiopods, arthropods, chordates, and echinoderms all post-date the beginning of the period by 10 to 25 million years. Trace fossils suggest earlier appearances for some groups, notably arthropods, but the observation remains that the Early Cambrian contains considerable time for the assembly and diversification of crown group morphologies" . |
page uploaded 15 May 2002
unless otherwise indicated, content © Chris Clowes 2002 (last updated: 3 Mar 2002)
revised ATW050906, edited RFVS111203
checked ATW051008




 Image
of fossil and life reconstruction of Microdictyon.
Reconstruction from
Image
of fossil and life reconstruction of Microdictyon.
Reconstruction from