| Palaeos | Chlorobionta | |
| Plants | Chlorobionta |
| Page Back | Unit Back | Plants | Dendrogram | References | Taxon Index | Pieces |
| Page Next | Unit Next | Unit Home | Glossary | Time |
LIFE |--Eubacteria `--Eukarya |--+--Rhodophyta | `--CHLOROBIONTA | |--Chlorophyta | `--Charophyta | |--(various green algae) | `--Embryophyta | |--Bryophyta | `--Tracheophyta (rhyniophytes) | |--Lycophyta | `--Euphyllophyta | |--Monilophyta | `--Spermatophytata | |--trimerophytes | `--Spermatopsida `--+--Fungi `--Metazoa |--Chordata `--Protostomia |
Lists Glossary Taxa References "The Wearing of the Green" Evolution of Land Plants Green Algae Plants Conquer the Land The Devonian Period The Carboniferous Period The Diversity of Plants Chlorobionta (Prasinophyta) Chlorophyta Charophyta Embryophyta Bryophyta Tracheophyta Lycophyta Euphyllophyta Monilophyta Spermatophytata Links |
 Beginning in the
Archean era,
Cyanobacteria evolved photosynthesis, which enabled them to use sunlight
to draw carbon dioxide from the atmosphere and convert it to oxygen,
water and glucose (a simple carbohydrate). These could be considered
the first simple "plants" Plants therefore might be seen as any
organism that is able to use sunlight, carbon dioxide, and water, to manufacture
its own food, that is, as a special class of autotroph. However, that's
far too broad. It would include all kinds of things like diatoms,
chromists, and photosynthetic bacteria which have nothing to do with plants in a
phylogenetic sense. They are, to be sure, all within the subject
matter of a General Botany class. All of these groups share some essential
biochemistry. However, what they don't share is a common ancestor
to the exclusion of all other organisms. This similarity arises from (a)
convergent evolution and (b) the exchange of plastids.
Beginning in the
Archean era,
Cyanobacteria evolved photosynthesis, which enabled them to use sunlight
to draw carbon dioxide from the atmosphere and convert it to oxygen,
water and glucose (a simple carbohydrate). These could be considered
the first simple "plants" Plants therefore might be seen as any
organism that is able to use sunlight, carbon dioxide, and water, to manufacture
its own food, that is, as a special class of autotroph. However, that's
far too broad. It would include all kinds of things like diatoms,
chromists, and photosynthetic bacteria which have nothing to do with plants in a
phylogenetic sense. They are, to be sure, all within the subject
matter of a General Botany class. All of these groups share some essential
biochemistry. However, what they don't share is a common ancestor
to the exclusion of all other organisms. This similarity arises from (a)
convergent evolution and (b) the exchange of plastids.
The description above also fails because it is only partially correct, even as a general description. Plants not only breathe out (respire) oxygen, but parts of their tissues also respire carbon dioxide, just as animals (heterotrophs) do. These processes provide the plant with energy for growing and maintaining its life support systems, and go on at all times. During the sunlit day, more carbon dioxide is consumed than is released in respiration, but at night photosynthesis ceases and the plant respires only carbon dioxide, returning a portion of its carbon to the atmosphere.
One better approach to defining "plants" is the "Chlorobionta" hypothesis, as used on the Tree of Life site:
There are two major lineages of green plants. One consists of most of what have been classically considered "green algae" -- mostly microscopic freshwater forms and large seaweeds. The other lineage contains several groups of "green algae" that are more closely related to land plants. Because these two lineages are monophyletic, they have been placed in a single monophyletic group called green plants, or, in technical parlance, the subkingdom Chlorobionta ...
This suffers only from being vague. Is there
anything else in the box besides green algae and land plants? The
ToL authors don't suggest any other content. Alternatively, this  might be an attempt to
suggest a crown group: "the last common ancestor of
Chlorophyceae and evergreens and all of its descendants," or something
like that. That sounds like a workable definition, but that can't be right, since they include the
prasinophytes among the Plants. Some, but not all, prasinophytes would
be excluded from the plants by a crown group definition. We think what the
ToL authors actually had in mind is
an even better choice: the stem group "green algae > red
algae." This
includes all of the prasinophytes, all other green algae and all plants, as
those terms are normally used, but not much of anything else.
might be an attempt to
suggest a crown group: "the last common ancestor of
Chlorophyceae and evergreens and all of its descendants," or something
like that. That sounds like a workable definition, but that can't be right, since they include the
prasinophytes among the Plants. Some, but not all, prasinophytes would
be excluded from the plants by a crown group definition. We think what the
ToL authors actually had in mind is
an even better choice: the stem group "green algae > red
algae." This
includes all of the prasinophytes, all other green algae and all plants, as
those terms are normally used, but not much of anything else.
Why do we care about definitions? The price of admission to doing good science taking an explicit position, so that others can prove you wrong. A vague definition, such as ToL's original formulation, is not good science. Unless we know precisely what they mean by "plant" we can't really make testable statements about what are or are not plants, nor about what characteristics plants have or do not have, nor about whence they might have derived their characteristics. Without really crisp definitions, these issues quickly get bogged down in semantics and arm-waving. Arguably that is exactly what happened to the whole business of taxonomy for the better part of a century.
Of course, definitions can never be "wrong," in a
logical sense. However, they can be useless, if they fail to draw lines
within our area of  interest. A vague definition is always useless
because it draws no line at all. Phylogenetic definitions have revived the
whole business of evolutionary systematics because they are quite precise
and refer to historical events (e.g., the evolution of red and green
algae from a common ancestor), rather than to some man-made list of
(sometimes fuzzy) characteristics. However, this precision also comes at a price. A
phylogenetic definition is built around a phylogenetic hypothesis. Unlike
a definition, a hypothesis can be wrong. If so, any definition based on that
hypothesis usually must be abandoned, and a lot of good work may go down the
tubes.
interest. A vague definition is always useless
because it draws no line at all. Phylogenetic definitions have revived the
whole business of evolutionary systematics because they are quite precise
and refer to historical events (e.g., the evolution of red and green
algae from a common ancestor), rather than to some man-made list of
(sometimes fuzzy) characteristics. However, this precision also comes at a price. A
phylogenetic definition is built around a phylogenetic hypothesis. Unlike
a definition, a hypothesis can be wrong. If so, any definition based on that
hypothesis usually must be abandoned, and a lot of good work may go down the
tubes.
Suppose for example, that we interested in the evolution of birds. Our hypothesis is that birds are the sister group of dinosaurs, and that some "dinobird" was their last common ancestor. We thus define birds as Struthio (ostrich) > Struthiomimus (a theropod dinosaur which looked like an ostrich) and dinosaurs as Struthiomimus > Struthio. Sadly, after years of frustrating labor sorting out the characteristics of the supposed dinobird ancestor, we realize that birds are dinosaurs. Oops. Our definition of "bird" turns out to include embarrassingly unbirdlike things like therizinosaurs, while our definition of "dinosaur" includes only tyrannosaurids and ornithomimosaurs. How to explain this little faux pas to those notoriously humorless folk whose grants supported our research the last three years? Again, that is simply the price of doing good science.
For that reason, we should be careful, as well as explicit, in framing the definition and articulating the underlying hypothesis. Here, the hypothesis is that red algae, in a colloquial sense, are closely related to green plants, in an equally colloquial sense. This then allows us to define both rigorously in terms of that relationship. Strictly speaking, we should do so in terms of particular anchor taxa, just in case either group turns out to be polyphyletic (which is possible). By all means, then, let's do so. On the red algae side, we'll pick Gelidium coulteri, a randomly chosen species of a well-known and very successful genus of red algae. On the green plant side, let's use an angiosperm, a highly derived group, and Quercus albus, because (as any citizen of the state of Connecticut will know) it symbolizes the willingness to take risks to vindicate historical truth. Based on our phylogenetic hypothesis, our working definitions are Chlorobionta (plants) = Q. alba > G. coulteri, and Rhodophyta (red algae) = G. coulteri > Q. alba.
Was that so hard? Of course not. But then, unlike ToL, we are not subject to the temptations to waffle which come with peer review and the caprice of granting agencies. Lest we be misunderstood, we support both peer review and post hoc review by grantors as excellent things for science; but they are not unmixed blessings. The inducements to please everyone may become irresistable. Now, unlike ToL, the purpose of Palaeos is only to amuse those who write it. However, if we can, occasionally, counterweight the temptation for others to hide behind intentionally vague and inconsistent pronouncements made in the service of their own comfort, perhaps it may serve another purpose as well.
ATW041201. Text public domain. No rights reserved.
The Green Algae - the Chlorophyta and Charophyta - include a number of mostly aquatic forms, including some unicellauar and primitive colonial forms. and other multi-cellular types that however lack a true root system
They are very closely related to (and probably the ancestors of) the higher plants in the kingdom Plantae. Molecular and cellular similarities between green algae, particularly the charophytes, and land plants include the following:
(1) Both the green algae and plants have chlorophyll b and beta-carotene
(2) Green algae and plants both have special intracellular membranes (the thylakoid membranes) which contain the chlorophyll stacked into grana.
(3) Charophytes have a cellulose content of 20 to 25% of the cell wall, a composition similar to that of plants.
(4) Cell division in green algae is very similar to that of land plants. Both use microtubules to bring vesicles containing new material in to form the cell plate which will divide the cell into two.
(5) Nuclear genes and RNA are similar between charophytes and plants.
 If the great evolutionary radiation of metazoa (multicellular animals) in the earliest Cambrian
oceans was the first great dramatic even of the Phanerozoic era (indeed ushering in the Phanerozoic), the conquest of land by
multicellular plants was the next, and of equal importance. Indeed, without the plants no animals would ever have been able to survive on land.
If the great evolutionary radiation of metazoa (multicellular animals) in the earliest Cambrian
oceans was the first great dramatic even of the Phanerozoic era (indeed ushering in the Phanerozoic), the conquest of land by
multicellular plants was the next, and of equal importance. Indeed, without the plants no animals would ever have been able to survive on land.
But whereas the Cambrian explosion was very rapid, in the order of perhaps 3 to 5 million years for the origin of all major phyla (and many others now extinct), the colonization of the land by vegetation was a much slower and more protracted. The reason for this is not hard to understand. Cambrian animals were moving into a favorable new environment with no competitors. Plants had to brave desiccation, extremes of temperature, and harsh ultra-violet radiation.
Enigmatic traces are known from the early and middle Ordovician, These are fossils of spores, cuticles, and tubes and don't reveal much about the structures or nature of these plants. All we can say is that these plants were probably of a hepatophyte grade of evolution - small, non-vascular, and lacking morphological differentiation into roots, stems, and leaves, like modern mosses and liverworts.
The first unambiguous record of land plants is from the Silurian period. They were mostly small, primitive forms, dependent on the proximity of water, and with the most rudimentary stem and leaf structure.
A common middle Silurian to early Devonian plant is Cooksonia, which had dichotomous branching and terminal sporangia (spore cases) at the tips of its green leafless stems. It is not known whether Cooksonia was a proper vascular (tracheid-bearing) plant. True vascular plants evolved and began to diversify during the Latest Silurian and Early Devonian.
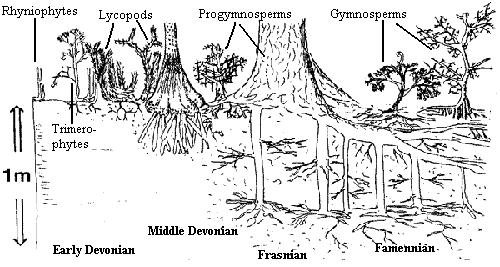 The Devonian period. marked a major shift in plant evolution and terrestrial ecosystems. Early Devonian plants such as the rhyniophytes,
zosterophyllophytes and lycopsids have features such as
vascular
tissue, stomata, a cuticle to
protect against drying, rhizoids, and sporangia at the tips of short lateral branches instead of terminal as in Cooksonia. These forms were small, non-rooted or shallowly
rooted, lacked woody tissue and hence were unable to grow beyond the height of small bushes. These plants reproduced by means of spores, which requires a moist habitat. They were therefore confined to moist,
lowland habitats, thus having little effect on their physical environment
The Devonian period. marked a major shift in plant evolution and terrestrial ecosystems. Early Devonian plants such as the rhyniophytes,
zosterophyllophytes and lycopsids have features such as
vascular
tissue, stomata, a cuticle to
protect against drying, rhizoids, and sporangia at the tips of short lateral branches instead of terminal as in Cooksonia. These forms were small, non-rooted or shallowly
rooted, lacked woody tissue and hence were unable to grow beyond the height of small bushes. These plants reproduced by means of spores, which requires a moist habitat. They were therefore confined to moist,
lowland habitats, thus having little effect on their physical environment
The first shrub and tree-like plants, such as Progymnosperms and lycopsids, had evolved by the middle Devonian. By the late Devonian the first real trees, such as Archaeopteris ("ancient fern" - not to be confused with Archaeopteryx, "ancient wing", the first bird!), had appeared. Trees have special vascular systems to allow for water circulation and nutrient flow against the pull of gravity. At the very end of the Devonian seed-bearing (gymnosperm) plants appeared for the first time, breaking free of the dependence on moisture that limits spore-bearing (pteridophyte) plants. Along with these developments came the development of advanced root systems and the production of soils, increased weathering, and huge ecological feedback.
The black & white figure shows the increasing terrestrial plant root depth penetration with time during the Devonian, leading to increasing soil depth and weathering. "Rhyniophytes" are a basal radiation of land plants such as Aglaophyton or Horneophyton. Trimerophytes include such plants as Psilophyton. Large lycopsids arrived in the Middle Devoinian. They originally appeared as low-lying herbaceous forms, such as Asteroxylon or Drepanophycus. Tree-sized lycopsids (e.g., Lepidosigillaria and Cyclostigma) appeared by the end of the Middle Devonian. Progymnosperms, such as Tetraxylopteris, arose in the Frasnian. By the Famennian, Archaeopteris forests are common. At the very end of the Devonian, Archaeopteris is found together with early gymnosperms, such as Elkinsia and Moresnetia, and zygopierid ferns such as Rhacophyton.
Image: from Prehistoric Animals, J. Augusta, illust. by Z. Burian, (Paul Hamilyn, London, 1960), pl.5, and Life Before Man by Zdenek V. Spinar, illustrated by Zdenek Burian
 The
Carboniferous Period
The
Carboniferous PeriodDespite the origin of the seed habit, the majority of Carboniferous plants reproduced by spores. The moist swampy environments of the time provided a nurturing environment. Lycophytes (scale trees and club mosses), which had evolved as small plants during the late Silurian? or early Devonian, and diversified greatly during the succeeding Devonian period, continued and thrived throughout the Carboniferous, but being dependent on water and moist conditions, most died out with the increasing aridity at the end of the Paleozoic, only a few small ones making it through. Calamites and ferns were other spore-bearing plants that appeared during the Devonian and flourished during the following Carboniferous period.
Image: Psaronius, a common Late Pennsylvanian tree fern, image from
Earth History Resources (former site).
| Page Back | Page Top | Unit Home | Page Next |
text by M.Alan Kazlev Creative Commons Attribution, except text signed ATW which is public domain
page uploaded 7 April 2002, last modified ATW090227, MAK120215; checked ATW050515