| Palaeos: Mesozoic | Mesozoic era | |
| Mesozoic Era | Mesozoic - 2 |
| Page Back | Back: Paleozoic | Back: Neoproterozoic | Up: Mesozoic - 1 | Unit Home |
| Page Next | Next: Cenozoic | Down: Triassic | Timescale |
| Palaeos: Mesozoic | Mesozoic era | |
| Mesozoic Era | Mesozoic - 2 |
| Page Back | Back: Paleozoic | Back: Neoproterozoic | Up: Mesozoic - 1 | Unit Home |
| Page Next | Next: Cenozoic | Down: Triassic | Timescale |
Mesozoic oceans were populated by a rich and diverse fauna of fish, reptiles, and a variety of cephalopod mollusks including the ammonites and the belemnites (nautiloids were also present but less common). Both these molluscan groups were adapted for speed and mobility. Fish were mostly slow moving heavy scaled ("ganoid") types, which were probably not as agile (teleosts only become predominant towards the later Cretaceous).
From the Jurassic onwards Plankton increased in diversity, with phytoplankton such as coccoliths, diatoms and silicoflagellates together with a zooplankton dominated by foraminifera and radiolarians. Arthropods such as the amphipod, decapod and isopod crustaceans together with nudibranch mollusks and the annelid and polychaete worms were also probably part of the plankton. MAK020428
The fossil record of Mesozoic annelids, like the fossil record of all annelids, is poor. We can only make a few, general remarks.
The end-Permian extinction more or less destroyed the entire Paleozoic benthic fauna. The Mesozoic benthic communities, developed an entirely new style, possibly (i.e., this is complete speculation) based on the very few anoxia-tolerant detritivores who would have flourished in the benthic carnage of the end-Permian. Whatever their origin, Mesozoic and Cenozoic benthic communities are dominated by infaunal (burrowing) deposit-feeders, rather than epifaunal suspension feeders. This was surely good for the annelids who are quite handy with low-oxygen, burrowing ways of making a living. Oligochaetes probably evolved in the Late Jurassic. However, they were unable to employ the usual annelid skills on land until the Late Cretaceous, when angiosperms began creating large quantities of humus, permitting the evolution of the oligochaete earthworms.
Brachiopods suffered greatly during the end-Permian extinction. They were able to make a considerable come-back during the Late Triassic, but ultimately declined and were ecologically replaced by bivalves. Their fate may have been tied to substrate. The brachiopods of the Late Triassic resurgence were strongly associated with carbonate shelves, the classic reef environment of the Late Paleozoic and Early Mesozoic. The rise in sea levels during the Jurassic and Early Cretaceous drowned these platforms on a global basis. That is, the residents of the carbonate platforms gradually found themselves too deep in the water column for sunlight to sustain photosynthesis, and the shelf ecosystems collapsed. This permitted the bivalves to "mussel" their way in, as they were better adapted to the soft and unstable sand & mud sea bottoms within the new photic zone. In fact, with the evolution of the rudists, the bivalves were able to make their own quick and sloppy reefs on even the softest substrate.
As a consequence, the surviving Mesozoic brachiopods became off-shore specialists, occupying deeper-water and more cryptic environments in crevices and on submarine cliffs below the photic zone. Some developed poisonous tissues. The more robust and globose terebratulides such as Terebratella and probably some species of Tichosina were free on the substrate. A few of these developed semi-infaunal strategies.
Mesozoic brachiopods, like many other invertebrates, show considerable differentiation between Tethyan (tropical) and Boreal (subtropical and temperate) types in the Late Triassic and Jurassic. Also like many other invertebrates, these distinctions broke down in the Cretaceous, as rising sea levels and flattened climate zonation homogenized most marine fauna.
Early Mesozoic bryozoans were largely cheilostomes and cyclostomes. During the Early Cretaceous, however, the cyclostomes declined while cheilostomes diversified. The reasons for this replacement are unclear. Both suffered massive extinctions in Maastrichtian time, possibly coinciding with the more general KT extinctions. The cheilostomes rebounded during the Cenozoic. The cyclostomes generally did not. McKinney & Taylor (2001).
 Mesozoic
cnidarians are mostly known from their greatest success story, the scleractinian
corals. Several groups of scleractinians developed tight symbiotic
relationships with photosynthetic zooxanthellae with a resulting huge boost to
their productivity. The scleractinians suffered considerably from the
drowning of the carbonate platforms on which their reefs were based during the
Late Jurassic and Cretaceous. However, they recovered quickly after the KT
extinctions.
Mesozoic
cnidarians are mostly known from their greatest success story, the scleractinian
corals. Several groups of scleractinians developed tight symbiotic
relationships with photosynthetic zooxanthellae with a resulting huge boost to
their productivity. The scleractinians suffered considerably from the
drowning of the carbonate platforms on which their reefs were based during the
Late Jurassic and Cretaceous. However, they recovered quickly after the KT
extinctions.
The End-Permian extinction at the end of the Paleozoic Era took a heavy toll on the stemmed echinoderms. The blastoids became extinct at that time and the crinoids suffered heavy losses. In general, Paleozoic echinoderms were epifaunal suspension and detritus feeders. Like so many high school students, their strategy was to sit more or less stationary on the sea bottom with their mouths open and wait for food to come to them. In the Mesozoic and Cenozoic, the echinoderms became more like undergraduates -- still bottom-feeders, but now willing to dig for it (infaunal detritus feeders) or, if sufficiently pressed, to go and hunt for it (armored herbivores and carnivores).
 This use of rather heavy armor runs counter to a general trend among Mesozoic
life forms to shed heavy plates and to depend more on speed, or on other
behavioral adaptations for survival. However, behavioral strategies depend
on having the neural equipment to select a response and adapt it to local
conditions. Echinoderms are poorly adapted for this sort of thing because
they are attractive, but brainless. So as time went on, echinoderms, like other
attractive but brainless organisms, were increasingly forced to rely on heavy
make-up, intimidating ornament, and a thick skin. The surviving crinoids,
for example, were articulates, with rounded, closely fitting armor plates,
usually bearing elaborate ornamentation. Some also gave up sessile life,
left their stems behind, and became motile. These swimming crinoids, the
Rovecrinidae, are discussed briefly elsewhere.
This use of rather heavy armor runs counter to a general trend among Mesozoic
life forms to shed heavy plates and to depend more on speed, or on other
behavioral adaptations for survival. However, behavioral strategies depend
on having the neural equipment to select a response and adapt it to local
conditions. Echinoderms are poorly adapted for this sort of thing because
they are attractive, but brainless. So as time went on, echinoderms, like other
attractive but brainless organisms, were increasingly forced to rely on heavy
make-up, intimidating ornament, and a thick skin. The surviving crinoids,
for example, were articulates, with rounded, closely fitting armor plates,
usually bearing elaborate ornamentation. Some also gave up sessile life,
left their stems behind, and became motile. These swimming crinoids, the
Rovecrinidae, are discussed briefly elsewhere.
However, for the most part, the old crinoid fauna simply died out. The future of the Echinodermata lay with the Echinoidea and Asteroidea. Echinoids are rare in Paleozoic faunas, but radiated extensively during the Mesozoic and Paleogene. Paleozoic, and even Triassic, urchins have no compound plates, and the interambulacral plates are constructed in many columns [1]. These earliest sea urchins are generally small and lack strong spine development -- characters which developed over the course of the Mesozoic.
Sponges as a whole did well and slowly diversified until the very end of the Mesozoic. However, this general trend is made up of varying fates of different groups of sponges. Demosponges and calcisponges recovered from the end-Permian extinction and dominated the reef fauna once more in many locations during the Late Triassic. However, they were gradually replaced by scleractinian corals. Hexactinnelids and some stromatoporoids continued as important frame builders for the coral reefs of Jurassic Europe. Demosponges and hyalosponges became more common in the Cretaceous. As sea levels rose, these sponges were sometimes able to thrive in regions which had become too deep for the corals. Mesozoic stromatoporoids (demosponges probably not related to the Paleozoic forms) were significant reef-builders in the Cretaceous. All types of reef-building sponges virtually disappeared at the KT boundary and never recovered. ATW040905
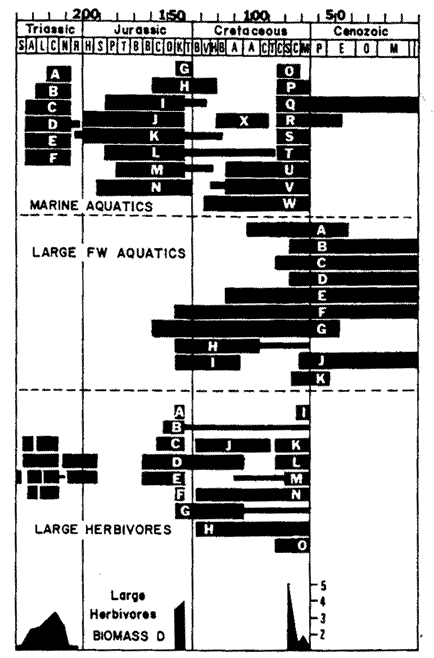 The Mesozoic era was an extremely long period of time, which saw
the rise and fall of successive "dynasties" of life.
At least half a dozen succesive evolutionary communities or empires of land vertebrates (tetrapods) can be distinguished.
Identifying them by characteristic large herbivores, these can be called the lystrosaur (Earliest Triassic [Induan]),
kannemeyeriid- traversodontid (primarily Gondwanan, though this may be sampling bias) (Early [
Olenekian] to Late (Carnian) Triassic ], plateosaur-vulcanodontid (Late Triassic
[Norian] -
Early Jurassic), sauropod- stegosaur (Middle to Late Jurassic), iguanodont-
nodosaur (Early to Mid Cretaceous), and ceratopsian-hadrosaur
(Late Cretaceous - Laurasia only, Gondwana was predominantly Titanosaurid, with Abelisaurid carnivores) communities or "empires".
In the sea one finds what could be perhaps termed the mixosaur- nothosaur (Mid Triassic), shastasaur (Late Triassic),
ichthyosaur- plesiosaurid- rhomaleosaurid
(Latest Triassic [Rhaetian] - Early Jurassic), ophthalmosaur- pliosaurid-
metriorhynchid (Middle Jurassic-Early Cretaceous),
and protostegid- elasmosaurid- mosasaur communities (Mid to Late Cretaceous).
The Mesozoic era was an extremely long period of time, which saw
the rise and fall of successive "dynasties" of life.
At least half a dozen succesive evolutionary communities or empires of land vertebrates (tetrapods) can be distinguished.
Identifying them by characteristic large herbivores, these can be called the lystrosaur (Earliest Triassic [Induan]),
kannemeyeriid- traversodontid (primarily Gondwanan, though this may be sampling bias) (Early [
Olenekian] to Late (Carnian) Triassic ], plateosaur-vulcanodontid (Late Triassic
[Norian] -
Early Jurassic), sauropod- stegosaur (Middle to Late Jurassic), iguanodont-
nodosaur (Early to Mid Cretaceous), and ceratopsian-hadrosaur
(Late Cretaceous - Laurasia only, Gondwana was predominantly Titanosaurid, with Abelisaurid carnivores) communities or "empires".
In the sea one finds what could be perhaps termed the mixosaur- nothosaur (Mid Triassic), shastasaur (Late Triassic),
ichthyosaur- plesiosaurid- rhomaleosaurid
(Latest Triassic [Rhaetian] - Early Jurassic), ophthalmosaur- pliosaurid-
metriorhynchid (Middle Jurassic-Early Cretaceous),
and protostegid- elasmosaurid- mosasaur communities (Mid to Late Cretaceous).
The following diagram is from Fig. 3. of Dr Robert T. Bakker's 1977 paper "Tetrapod Mass Extinctions - A model of the regulation of speciation rates and immigration by cycles of topographic diversity" in A. Hallam, ed. Patterns of Evolution as illustrated by the Fossil Record, Elsevier Scientific Publishing Company, Amsterdam, Oxford, New York, pp.439-68
Although subsequent research has modified some of the family rankings and stratigraphic correlations, the basic pattern remains.

During the Mesozoic era there were a number of lineages of marine reptiles. In comparison with the mammals, many more types of reptiles, having attained an existence on land, returned to the seas. Aquatic adaptation (whether freshwater, estuarine, or marine) is a common phenomenon among reptiles, due to their low metabolic rate, tolerance of anoxia and of low body temperatures, and easy ability to make use of fermentative metabolism for muscle activity. Moreover it does not require great structural or physiological changes, as indicated by the modern marine iguana. Reptiles move with a naturally sinuous motion, which is easily adaptable to swimming (as it comes originally from the swimming motion of the fish). In marine iguanas aquatic locomotion requires only one quarter the metabolic activity of terrestrial locomotion. When looked at this way, it is not surprising that reptiles have returned to the water, like their tetrapod and fish ancestors, whenever conditions were favourable.
The following is a list of aquatic reptiles, with some basic data. Note: Many of these pages are under construction or very incomplete. So here they are: the marvelous Mesozoic marine reptiles:
| Picture | name | time-span | habitat | location | approx size | food |
|
|
Plesiochelyidae | late Jurassic [Kimmeridgian] to early Cretaceous | estuarine, near shore marine | Central to East Laurasia | shell c.75 cm??? | invertebrates, fish, plant material? |

|
Desmatochelyidae + Protostegidae | Cretaceous
Desmatochelyidae - Albanian to Maastrichtian Protostegidae - Turonian to Maastrichtian |
open ocean | Desmatochelyidae - cosmopolitan
Protostegidae - west Laurasia ("inland sea") only? |
Desmatochelyidae - av. 1.2 meters (shell)
Protostegidae - up to 4 meters long (Archelon) |
jellyfish, other invertebrates, fish? |
|
Toxochelyidae | late Cretaceous (Coniacian to Maastrichtian) | open ocean | west Laurasia ("inland sea" to west Atlantic) | shell 60-120 cm | jellyfish, other invertebrates, fish? |
 |
Cheloniidae | latest Cretaceous [Maastrichtian] to Recent | open ocean | Worldwide | 75 cm to over 1 meter long | jellyfish, other invertebrates, fish? |
 |
Ichthyosauria | Triassic to Jurassic,
a few stragglers into the Cretaceous |
open ocean | world wide | 1 to 23 meters | mostly fish, also cephalopods, smaller reptiles |
 |
Placodontia | Triassic | near shore marine | Tethys Sea | 1 to 2.5 meters | probably shellfish (Bivalve mollusks) |
| Pachypleurosauridae | Middle Triassic [Early Anisian to Ladinian] | estuarine, near shore marine | world wide | 20 cm to 1 meter | fish, crustacea, etc | |
 |
Nothosauridae | Middle to early late Triassic [Anisian to Carnian] | near shore marine | world-wide | 2 to 8 meters | mostly fish |
 |
Plesiosauria | rare in the Triassic
common in the Jurassic and Cretaceous |
open ocean
( a few estuarine species) |
world wide | 2 to 14 meters | fish, cephalopods, other reptiles |
 |
Mosasauroidea | Aigialosauridae Late Jurassic to Early late Cretaceous (Tithonian to Turonian)
Mosasauridae: late Cretaceous (Turonian to Maastrichtian) |
open ocean | Aigialosauridae: Europe
Mosasauridae: world wide |
Aigialosauridae: av. 1 meter
Mosasauridae: 2.5 to 17 meters |
fish, cephalopods, other reptiles |
 |
Thalattosauria | Triassic | near shore marine | Tethys Sea, also west Laurasia | about 1.5 meters | fish, crustacea etc |
| Nanchangosaurus | earliest Middle Triassic (Early Anisian) | near-shore and estuarine | China | no info (~1 meter?) | probably fish, crustacea, etc | |
 |
Teleosauridae | Jurassic | estuarine, near shore marine | world wide | 2 to 6 meters | mostly fish |
| Metriorhynchidae | Jurassic to Early Cretaceous | open ocean | world wide | 2 to 6 meters | mostly fish |
MAK020428, modified MAK030915
