| Palaeos | Choristodera | |
| Vertebrates | Neochoristodera |
| Page Back | Unit Home | Unit Dendrogram | Unit References | Taxon Index | Page Next |
| Unit Back | Vertebrates Home | Vertebrate Dendrograms | Vertebrate References | Glossary | Unit Next |
|
Abbreviated Dendrogram
DIAPSIDA |?-CHORISTODERA | |--Cteniogenys | `--+--Monjurosuchus | `--Neochoristodera | |--Simoedosauridae | `--Champsosauridae |--ARCHOSAUROMORPHA `--LEPIDOSAUROMORPHA |
Contents
|
The phylogenetic position of the Choristodera generally is still fairly vague. Current estimates place them somewhere between basal Diapsida and basal Archosauromorpha. (In fact, one well-known web site has them in both places) Here, they are placed as the sister group to Archosauromorpha, consistent with deBraga & Rieppel (1997). This paper does seem to contain some probably minor errors in scoring the Choristodera. For example, the pterygoid flange is scored as having a single row of teeth along the posterior margin, rather than a shagreen of small denticles, as might be more appropriate. It is unlikely that these significantly affect the result. However, it remains entirely possible that the Choristodera are more nearly basal diapsids than near - Archosauromorphs. Internal relationships within Choristodera are described according to Gao & Fox (1998). The Choristodera in general, and particularly the Champsosauridae, are plagued with bad names -- junior synonyms of unrelated genera and the like. Gao & Fox do a very careful job of trying to straighten out this confusion, and their paper is strongly recommended.
The Champsosaurs are the "classic" Choristoderans: very crocodile-forms with highly elongated snouts, looking very much like living gharials. Much of the similarity is illusory. The orbits, for example, are placed well forward of the archosaur position. The rear of the skull is not flat and strongly consolidated, like a crocodile. Instead it is rounded, hugely expanded and consists to a considerable degree of complex bony arches surrounding empty space. In life, this space was presumably occupied by massive jaw muscles. However, the fossils look a bit like a lizard skull in a carnival mirror, the anterior drawn out into a long, thin pisciverous rostrum and the posterior blown up like a balloon. However, champsosaurs lack the complex quadrate of lizards as much as they lack typical archosaurian features.
One of the striking results of deBraga & Rieppel's study is the number of presumably convergent features the choristoderes share with placodonts. Some examples (all apomorphies of both groups) include (a) the transverse process of the pterygoid extends below the maxillary tooth row; (b) the mandibular joint lies posterior to the level of the occiput; (c) the maxilla does not extend to the level of the posterior margin of the orbit; and (d) the post-orbital skull is longer than the pre-orbital skull. One might suspect that these odd convergences in jaw structure are derived from a common lifestyle. Specifically, it seems quite possible that the choristoderes were originally shellfish-eaters like placodonts and only secondarily became long-snouted fish eaters. Interestingly, choristodere families more primitive than champsosaurs do tend to have shorter, broader snouts.
The limbs are very well developed for basically aquatic animals. It appears from their architecture that the entire distal limb, not just the hands and feet, was used as a paddle. This is also consistent with the relatively short limbs that may have allowed these animals some mobility on land, and the straight, unbowed ulna. The limbs are relatively gracile. Champsosaurs would not have been ambush amphibious predators like crocs since they seem to have lacked the ability to leap to attack in the amazing fashion of modern crocodiles, who can often jump 1.5 times their length. In addition, champsosaur ribs are thickened and short in the manner of other aquatic reptiles, such as some pachypleurosaurs.
Choristoderans do share one other feature with crocs. They survived the KT boundary. They became extinct only in the Eocene, possibly even the Oligocene. Their ultimate extinction may have been the result of any number of factors: falling temperatures and the evolution of whales come to mind. In this connection, it may be relevant that their last redoubt may have been the non-marine waters abutting the cooling remnants of the Western Interior Sea, rather than the whale-infested waters of the Tethys and Atlantic. ATW000716.
Range: Middle Jurassic to Eocene.
Phylogeny: Choristodera : Cteniogenys + (Hyphalosaurus + Monjurosuchus + * : Simeodosauridae + Champsosauridae)
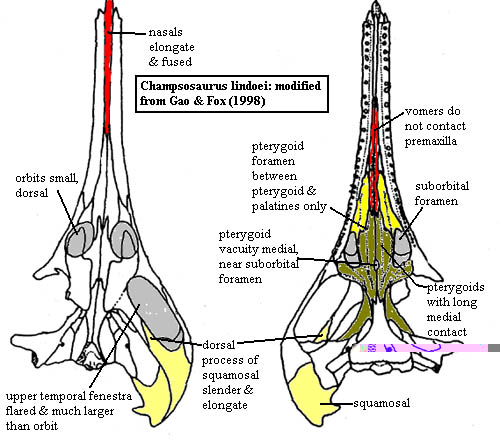
Characters: $ Basal infolding of tooth enamel; $ vomer does not contact premaxilla (snout elongated); $ palatal foramen (?) opens between pterygoid & palatine without contribution from vomer; $ lacrimal foramen opens between lacrimal, prefrontal and palatine -- not penetrating lacrimal; $ pterygoids with long medial contact; $ interpterygoid vacuity medial, small & near posterior margin of suborbital fenestra; $ orbits small & directed dorsally; nasals long and fused; $ upper temporal fenestra flared posteriorly and much larger than orbit; dorsal process of squamosal slender & elongate to mid-level of lower temporal fenestra; $ pterygoid process of quadrate horizontal, reduced & slender; $ presacral vertebral centra short & spool-like; $ "ventromedial crest" of dorsal vertebrae reduced or absent; bones pachyostotic in adults.
References: Gao & Fox (1998); Tarduno et al. (1998). ATW010715.
Phylogeny: Neochoristodera : Champsosauridae + *.
 Champsosauridae:
Champsosaurus.
Champsosauridae:
Champsosaurus.
Range: lw(?)K-lwEc.
Phylogeny: Neochoristodera : Simoedosauridae + *.
Characters: Crocodile-like aquatic forms. Single fused nostril at tip of greatly elongated snout, exceeding 1/2 of skull length; mandibular symphisis extended to over 1/2 length of tooth row; splenial strongly involved in symphisis; secondary palate for air-breathing; premaxilla does not contact vomer; small interpterygoid foramen; suborbital fenestra short & kidney-shaped; pterygoid fused to parasphenoid (?); craniomandibular joint anterior to occipital condyle; internarial present; interorbital width narrower than diameter of orbit; postorbital excluded from orbit; lacrimal reduced to small triangle; neomorphic bone forms ventral border of post-temporal fenestra; paroccipital process strongly deflected ventrally; temporal bars extended laterally to accommodate very large jaw muscles; limbs poorly ossified (but full development of all limbs). No sternum, but not relevant for aguilliform movement.
Links: Evolutionary Relationships of Archosaurs (basal archosauromorphs); Archosauria: More on Morphology; Theodore Roosevelt National Park Champsosaur; Welcome to FS-Today!; usda.gov article5; utoronto champsosaurus; champsosuarus gigas; tucsonshow tgms2000_p9; Introduction to the Diapsids; Champsosaur Bones in the Arctic; Brown Coulee - Champsosaur Vertebrae; Champsosaurs - Suite101.com; Ghost lineage; Fossil Find Above the Arctic Circle. ATW000531.
References: deBraga & Rieppel (1997); Gao& Fox (1998)
| Page Back | Unit Home | Page Top | Page Next |
checked ATW050908
Using this material. All material by ATW is public domain and may be freely used in any way (also any material jointly written by ATW and MAK). All material by MAK is licensed Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners