| Rostroconchia | ||
| Mollusca | Rostroconchia |
Conchifera
└─► |
|||
◄─┬─Conocardiida
└─Ribeirioida |
Mollusca |
|
Abbreviated Dendrogram
Mollusca
├─Aculifera
└─Conchifera
╞═Tryblidiida (Monoplacophora)
│ └─Cephalopoda
╘═Helcionelloida
├─Rostroconchia
│ ├─┬─Conocardiida
│ │ ╘═╤═Ribeirioida
│ │ └─Scaphopoda
│ └─Bivalvia
└─Gastropoda
|
Contents
Overview |
Taxa on This Page

image © xxxx from Lecture 8 Origin and Evolution of the Mollusca former page) / Lecture 11 Evolution of the Mollusca: bivalves (former page)
The Rostroconchia are a group of Palaeozoic bivalved molluscs that were originally considered Bivalves, but were then realised to belong to a distinct class of their own. Pojeta et al. 1972); Pojeta & Runnegar 1976). According to the latter workers, rostroconches are Conchifera with "an uncoiled and untorted univalved shell which straddles the dorsal midline, and a bivalved adult shell… [with] dorsal commissure lacking." They are characterized by a single, pseudobivalved shell which enclosed the mantle and muscular foot. The anterior, probably downwards-orientated, part of shell has a gape from which the foot may have emerged. They probably lived a sedentary semi-infaunal lifestyle
To date, about three dozen genera and a greater number of species have been described. Most rostroconches are small, often less than two centimeters in length, but large forms, about 15 cm long, can be found in Devonian limestones of the midcontinental United States.
The following sketch © xxxx from McRoberts 1998 shows the main parts of the rostrochonch shell.

Rostroconchs look superficially like bivalves and like them probably had an extensible foot, as indicated by a prominent anterior gape in the shell. However, the shape of the foot, and perhaps the internal anatomy, were probably much closer to the scaphopods, which, according to one theory, they gave rise to.
Rostroconchs began life as a small limpet-like, bilaterally symmetrical protoconch larva, probably (like the scaphopod larva) planktonic. From this the adult, likewise bilaterally symmetrical shell (dissoconch) grew down as a pair of valves.
The adult rostroconch superficially resembles a bivalve, but differs in that there is no functional hinge. Instead of a flexible ligament or articulating system that charcterises the bivalve hinge, some or all of the shell layers -- layers of stiff calcite -- are continuous across the region of the dorsal margin. The effect is that the rostroconch shell functions more like a taco shell than a clam shell. The two shell valves would have been rigidly fixed in place. Rather than having a hinge, the dorsal margin would, at most, have formed a poorly elastic structure. The shell apparently broke periodically at the margin to permit shell growth.
The posterior of the shell is usually elongated into a flattened tube called the rostrum through which inhalent and exhalent siphons may have emerged. It can be assumed that the animal positioned itself so that the rostrum was above the layers of sediment in which the creature had burrowed; perhaps also the rostrum which may have aided in water filtration.
The earliest known rostroconch genus, Heraultipegma, dates from the Late Terreneuvian (Tommotian), but is much more primitive than the typical or true rostroconchs. These appeared and flourished during the Early Ordovician, almost rivalling true bivalves, with which they may or may not have shared a similar lifestyle, in numbers and variety. But with the end-early Ordovician Floian) turn-over they declined, and only one order, the Conocardioida, continued until the Guadalupian (Middle Permian). Clarkson (1993); Paleobiology Database (acc. 080222).
The earlier and more primitive rostroconchs such as Ribeiroia (order Ribeirioida) had a hinge in which all the shell layers traversed the dorsal margin, resulting in a rigid shell. In more advanced forms, like Conocardium, the outer layer does not cross the margin, suggestiung an independent step towards the condition already achieved by bivalves. Conocardium, has a gape at one end and very pronounced rostrum at the other.
According to Engeser & Riedel 1997), during the Devonian, the conocardoid rostroconchs gave rise to the Scaphopoda, which would therefore be Rostroconchia (in the same way that birds are dinosaurs). MAK021208, revised ATW080222.
Rostroconchs almost certainly evolved from helcionelloid molluscs Pojeta & Runnegar, 1976; Peel, 1991; Wagner, no date) during the early Cambrian. It was originally suggested Runnegar & Pojeta, 1974; Pojeta & Runnegar, 1976) that they gave rise to the bivalves on the one hand (by separation of the valves and development of a proper hinge), and the Scaphopoda on the other (with the valves forming into a long tube). However, some cladistic and molecular evidence (Waller, 1998; Steiner & Dreyer, 2002) indicates that bivalves are only distantly related to scaphopods, but that cephalopods are closely related to scaphopods.
Embryological evidence (Scaphopod protoconchs) might support the thesis that Scaphopods evolved from rostroconchs. Engeser & Riedel (1997). Moreover, cephalopods (Pojeta, 1980) or both scaphopods and cephalopods Runnegar, 2002; Steiner & Dreyer, 2002), are closely related to, or descended from, laterally compressed helcionelloids. Now, these early primitive helcionelloids are precisely the same forms that can be seen as the ancestors of the Rostroconchia [Wagner no date]. So, the implication might be that helcionelloids gave rise to both rostroconchs and cephalopods, but not to Bivalves, and that rostroconchs in turn were the ancestors of scaphopods.
Truthfully, we find these ancestor-descendant hypotheses unnecessary and confusing. Like Steiner & Dreyer (2003), we reject them. Those authors argue that both molecules and morphology unite scaphopods and cephalopods. If so, then rostroconches are either some sort of probably) paraphyletic group at the base of the Conchifera, or they share a branch with the bivalves. The similarity of rostroconchs to the helcionelloids (Peel, 2004) is interesting, but doesn't help much, because the Helcionelloida are definitely) some sort of paraphyletic group at the base of the Conchifera. Therefore, the helcionelloid chracters shared by rostroconchs are likely to be uninformative characteristics of the ancestral conchiferans. Our choice is thus somewhat arbitrary, but it is natural to believe that the pseudo-bivalved rostroconchs might have a more than casual relationship with real bivalves; and that is where we have placed them. MAK021208, last revised ATW080222.
The Class Rostroconchia (Pojeta, Runnegar, Morris & Newell,1972) is usually divided into two orders, as follows:
Order Ribeirioida (Early Cambrian to Early Silurian)
Order Conocardiida (also spelt Conocardioida) (Furongian to Late Permian)
The Ribeirioida are actually a paraphyletic grade, as they give rise to the Conocardiida. The following cladogram by Wagner shows the relationships between the earliest Rostroconchia and their Helcionellid ancestors:
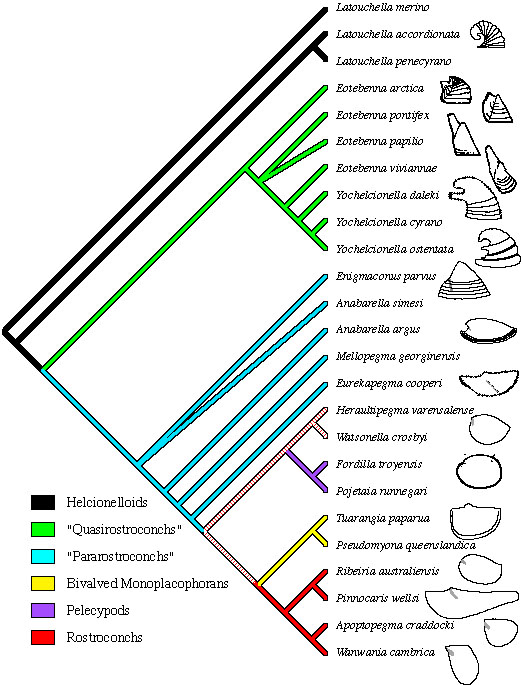
Note that this cladogram has the Bivalvia (Pelycopods - in purple) emerging from proto-rostroconchs. This disagrees with morphological cladistic evidence, according to which the Scaphopods are most closely related to Cephalopods, and only distantly to bivalves. Thus either Scaphopods or Bivalves, but not both, evolved from Rostroconch ancestors, in which case they independetly developed similar morphotlogies as a result of exploiting similar infaunal lifestyles, rather than actual ancestor-descent relationships. Some current molecular phylogenies however have scaphopods, bivalves, and gastropods forming a single clade (with gastropods either closer to bivalves or scaphopods). In this case simialrities between scaphopods and cephalopods are due to convergence. This hypothesis is hard to explain ecologically, as the two evolutionary lineages would have occupied very different ecological niches (infaunal and weakly nektonic repsectively). So either the current (circa 2011) molecular consensus is completely wrong, or proto-cephalopods also went through an infaunal stage. revised MAK120614
Range: Cambrian Epoch 2 to Late Permian
Phylogeny: Paraphyletic Helcionelloida : Gastropoda + Bivalvia + * : (Conocardiida + (Ribeirioida + Scaphopoda)
Pageby M. Alan Kazlev, uploaded 8 December 2002, revised ATW080222, checked ATW080220, last revised MAK120614 Creative Commons Attribution