| Palaeos | Chelonii | |
| Vertebrates | Centrocryptodira |
| Page Back | Unit Home | Unit Dendrogram | Unit References | Taxon Index | Page Next |
| Unit Back | Vertebrates Home | Vertebrate Dendrograms | Vertebrate References | Glossary | Unit Next |
|
Abbreviated Dendrogram
ANAPSIDA ? `--o CHELONII |--Odontochelys `--Testudinata `--+--Proganochelys `--+--Kayentachelys `--+--Meiolanoidea `--+--Pleurodira |--Paracryptodira `--+--Protostegidae `--+==Xinjiangchelyidae `--o Centrocryptodira |--+--Hoyasemys | `--Sinemydidae |--Osteopygis |--Trionychoidea `--o Durocryptodira |--+--Kirgizemys | `--+--+==o Macrobaenidae | | |--Judithemys | | `--Macrobaena | `--o Chelonioidea-Chelydridae-Kinosternoidea Clade | |--+--Chitracephalus | | `--Chelonioidea | `--+--Chelydridae | `--Kinosternoidea `--Testudinoidea |
Contents
Overview |
Taxa on This Page
The names Centrocryptodira and Durocryptodira nicely represent the conflict between morphology and molecules in turtle evolution; neither is used much now.
The morphology-based Hennigian cladistic taxon Centrocryptodira was named by Gaffhey and Meylan, 1988 to include Chelonians with pronounced cervical articulations, which includes the Meiolaniidae, the paraphyletic Sinemydidae-Macrobaenidae assemblage, and the Polycryptodira (Gaffney, 1996).
The Meiolaniidae have since been considered a stem Chelonian by more recent workers (Hirayama et al, 2000, Joyce 2007, Sterli 2008, and Anquetin 2009); this difference would seem to be due to the replacement of Hennigian cladistics with statistical-computational parsimony analysis, and phylogenetic nomenclature such as restricting morphology based definitions to crown-groups only (in this regard, Joyce's 2007 paper is as important and revolutionary as Gaffhey 1975 and Gaffhey and Meylan, 1988.
This more recent paradigm means the rejection of cladograms based on a smaller number of well-known characters with those based on a greater number of less analysed and atomistic treated characters. This shift in emphasis is itself the result of the development of cheaper and more powerful computing. For this reason, in every field of cladistic paleontology, the Hennigian "hand-coded" method has been replaced by trees based on huge matrixes and supermatrixes of the sort that could only be calculated by powerful computers.
Both methodologies agree however that the various Cretaceous to Paleocene semiaquatic and mostly Asiamerican turtles of the families Sinemydidae and Macrobaenidae are a paraphyletic assemblage or grade of forms more advanced than Jurassic Asian Xinjiangchelyidae, and from which recent or crown group cryptodires evolved. Here the Polycryptodira Gaffney and Meylan, 1988 are synonymous with "Crown Group Cryptodira" of statistical-computational and phylogeny-nomenclature based current cladism, and includes the superfamilies Chelydroidea, Chelonioidea, Trionychoidea and Testudinoidea.
For Gaffney and Meylan and co-workers and students therefore the series goes Xinjiangchelyidae + (Meiolaniidae + (paraphyletic Sinemydidae and Macrobaenidae + (Chelydroidea + (Chelonioidea + (Trionychoidea and Testudinoidea)))). While for later workers, it is Xinjiangchelyidae + (paraphyletic Sinemydidae and Macrobaenidae + (Chelonioidea + (Chelydroidea + (Trionychoidea and Testudinoidea)))). The only differences is that for the latter the Meiolaniidae are not cryptodires but placed outside the crown group testudines altogether, and Chelonioidea are the most basal taxon of crown group cryptodoires. Developmental biology supports the latter, with Chelonioidea more basal than Chelydroidea, and Trionychoidea and Testudinoidea the most derived Werneburg & Sánchez-Villagra 2009. But when matched with the fossil record there is very little congruence. Of advanced cryptodires only the trionychoidea goes back to the early-Late Jurassic, all the other clades are represented by extensive ghost lineages until at least the late Cretaceous
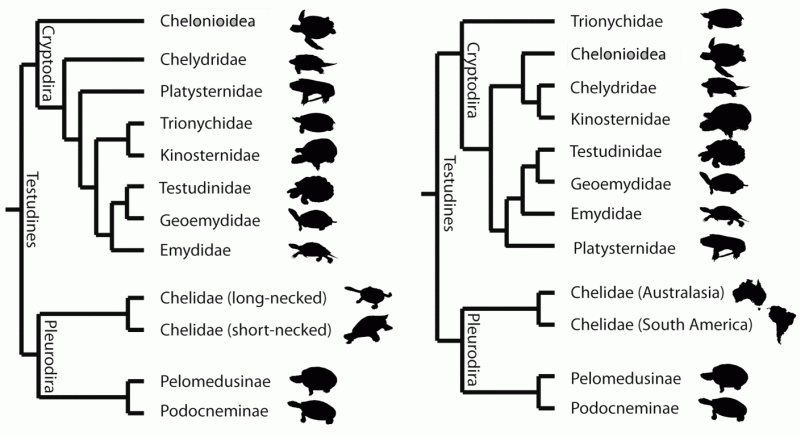 |
| Alternative testudine phylogenies. (A), morphology-based, according to Joyce 2007, with cheloniidea as the most basal extant cryptrodires. This is also supported by developmental biology (Werneburg & Sánchez-Villagra 2009). (B) molecular-based, according to Shaffer 2009, with Trionychia as the most basal extant clade, which is supported by the fossil record (Danilov & Parham, 2006). Diagram from Jones et al, 2012 fig 1; Creative Commons Attribution |
Molecular sequencing presents a totally different picture. Beginning with Shaffer et al. (1997) and ubiquitously verified by every paper since (e.g. Fujita et al, 2004, Krenz et al 2005, Shaffer, 2009, Barley et al. 2010, (more)). In contrast to morphological analyses, it finds both the Trionychoidea and Chelydroidea to be polyphyletic. In this phylogeny, the big headed turtle Platysternon megacephalum turns out to be more closely related to the Testudinoidea. Equally surprisingly, the traditional Trionychoidea splits into two groups, one of these, consisting of the families Trionychidae and Carettochelyidae (the Trionychia), being resolved as the most basal extant cryptodires, and only very distantly related to the other (the Kinosternoidea). What is surprising is that morphologically these are very specialised and similar animals; whether using Gaffney's Hennigian cladistics or the more popular current computational-statistical method of Joyce and later workers, the Trionychoidea ( as Trionychia + Kinosternoidea) is universally resolved as a highly derived monophyletic taxon.
The consensus molecular phylogeny therefore would be Pleurodira + (Trionychia + (Chelydridae + Chelonioidea + Kinosternoidea) + Testudinoidea incl. Platysternon). Even if the details of relation between the Chelydridae + Chelonioidea + Kinosternoidea, and of Testudinoidea and Platysternon (whether the latter is a sister group to the Testudinoidea as a whole or included within the group as the sister clade to Emydidae) might vary slightly, all agree on the fundamental position of the Trionychoidae at the base of the cryptodire family tree. In view of the great morphological similarity between the Trionychoidae and the Kinosternoidea, it seems mind-boggling that evolution should follow such identical nodes independently even after a hundred million years or more of seperate evolution.
Could the apparent basal position of the Trionychoidae then simply be an artifact of molecular sequencing? It is striking that the Trionychoidae have the longest branches of extant chelonians, which shows that their apparent basal position may be the result of sequencing artifacts, specifically the common problem known as long branch attraction. Sterli, (2010) tested the validity of the long branches of the Pleurodira as the adjacent group to Trionychoidae, with analyses excluding Pleurodira, Trionychoidae, and Kinosternoidea. The results show that the phylogenetic position of each clade is stable and does not vary when one is excluded from the analysis. As further confoirmation, the five genes were tested separately in Bayesian analyses and maximum likelihood. In all the cases the topologies retrieved Trionychoidae as the adjacent group of Pleurodira, as distinct from all other cryptodires (Sterli, (2010) p.103)
Support for the ancient status of the Trionychoidae comes from the stratigraphic sequence; an appearance as far back as the Oxfordian (early Late Jurassic) (Danilov & Parham, 2006) makes them the earliest known crown cryptodire lineage, the only extant clade whose unambiguous fossil record predates the late Cretaceous. Although many cladists dislike stratigraphy as it gets in the way of nice and tidy morphological analyses, the present author (MAK) considers excessive appeal to ghost lineages to be a cop out in the case of groups with an already extensive fossil record. Since even terrestrial testudines are easily and frequently fossilised, one needs to ask why so many clades are totally unknown as fossils for more than half their purported stratigraphic range.
Another intriguing piece of evidence is the presence of transitional forms, as well as exterme homoplasy. Basilochelys combines Xinjiangchelyid attributes with those of Adocusia. The mid Cretaceous proto-Carettochelyid Kizylkumemys has very archaic (paracryptodire-like) characteristics. Sandownia is another form that may be either a stem-eucryptodire-like trionychoidan or an unrelated animal convergent on trionychoidans. Cladistic analysis by Tong et al. 2009 reveals rampant homoplasy among testudines and several characters occuring only in Trionychoidae (Crown Trionychoidae, Adocus and Nanhsiungchelyidae) and some macrobaenidsinemydid turtles, which suggests the Trionychoidae might have a more stemward position among eucryptodiran turtles. This would explain both their very early stratigraphic occurrence in contrast to Kinosternoidae, Chelydroidea or Testudinoidea; and the results of molecular phylogenies (Tong et al. 2009 pp.168).
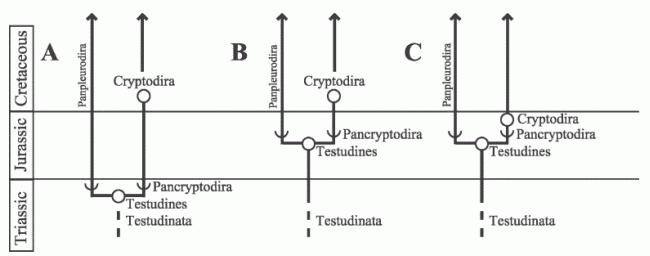 |
| Diagram shopwing three hypotheses of testudine evolution and the origins of the stem group or Pancryptodira and the crown group (here Cryptodira) of Cryptodire turtles. A, Traditional hypothesis (e.g., Gaffney and Meylan, 1988; Shaffer et al., 1997) which posits a very early split between Pleurodires and Cryptodires, and then long stem lineages with both; B, Stastistical-Parsimony cladistic methodology (e.g. Joyce 2004, Joyce 2007, ) which posts both a long stem testudine lineage and a long pancryptodire stem prior to diversgence opf crown groups; C, Molecular-Stratigraphic hypothesis which has a long stem-testudine lineage but a very early division of higher cryptodires (Trionychoidae + Durocryptrodira). as proposed by Danilov & Parham, 2006 and 2008. Taxa are stem based (open semi-circle) or node based (circle). Diagram from Danilov & Parham, 2006 p.578, fig 7 |
Based on a combination of paleontology and molecular phylogeny, Danilov & Parham, 2006 and 2008 propose a scenario in which Pleurodira, Trionychoidae, and non-trionychoidan cryptodires, the latter grouped together in the new clade Drurocryptodira (meaning hard shelled cryptodires, to distinguish them from the soft-shelled Trionychoidae), diverge during the middle to late Jurassic.
Both explanations cannot be correct. If hennigian morphological, statistical morphological, and developmental parsimony analyses are all to be preferred over stratigraphy and molecular evolution, Trionychoidae remarkably molecularly divergent from all other extant cryptodires of which all major groups must be represented by extended ghost lineages going deep into the Jurassic. That means that all other higher cryptrodire lineages unknown from fossils for about half their history, despite turtles otherwise having an excellent fossil record. This recalls the dilemma with Archaeopteryx and the dromaeosaurs. The late Jurassic Archaeopteryx is clearly more advanced than its early Cretaceous "ancestors". In this case late Jurassic ancestors like Anchiornis are known. Moreover, not only is the fossil record almost totally unreliable in recording the first appearance of even well-fossilised taxa, but molecular sequencing is also shown to have a very poor phylogentic signal, full of misleading artifacts such as basal Trionychoidae and Testudinoid Platysternids
But if molecular phylogeny is reliable, then so is stratigraphy, with the Pleurodires and the Trionychoidae well known from the late Jurassic, confirming their early branching in the molecular tree. Trionychans would be convergent with higher hard shelled turtles, especially the Kinosternoidea, the most extraordinary degree, much more so than protostegidae and dermochelyidae, since unlike protostegids, trionychoidans do not retain primitive features. This means that one after another unique adapation would have to occure independently in both lineages to arrive at morphologically convergent but phylogenetically distinct highly specialised forms; a bit like the humanoids of Star Trek, the vulcans, klingons, and so on who all independently evolved to almost identricxal human-like end forms (link). Morphologically this makes very little sense. It is one thing to imagine an ichthyosaur looking roughly like a dolphin, it is quite another for them to be so similar as to be included in the same superfamily, molecular phylogeny is full of convergences of this sort that don't make sense given the random nature of selection and evolution; another example are the Heliozoa (more). It also means that even molecular development follows molecular convergence, with very little useful phylogenetic signal. Similarily, molecular phylogeny recovers many morphological nonsensical clades, such as the Afrotheria, Cyclostomia, and archosaurian turtles
In the end we are left with two equally persuasive yet also equally inconclusive and problematic scenarios. Perhaps new fossil discoveries and new methodological insights can shed light on this enigma. There being no easy solution at present, we have included both phylogenies under each entry, MAK130126
The Sinemydids and Macrobaenids are a paraphyletic assemblage of basal eucryptodiran turtles of a more advanced (derived to use the technical term) grade than the Xinjiangchelyids (themselves most likely also a paraphyletic grade). In cladistic analyses they are usually resolved as successive sister taxa of Polycryptodira (Gaffney) or Crown Cryptodira (Joyce etc) ( Parham and Hutchison 2003, Gaffney et al. (2007), Joyce 2007, Rabi et al 2010), although some analyses seem to suggest grades or subgroups, although tree topologies differ according to whether one follows a Hennigian (e.g. Joyce) or a statistical (e.g. Joyce) cladistic methodology.
They include mostly terrapin-like semi-aquatic forms that first appeared during the Late Jurassic in China and are best known from the Cretaceous to Paleogene of Asia and North America. During the early Cretaceous, they seem to have outcompeted the similarily terrapin-like pleurodires (check ref, biogeography), who retreated to South America and Australasia. By the middle to late Cretaceous the Sinemydid-Macrobaenids had radiated into a number of distinct lineages; these became the Cenozoic testudine groups, the Chelonioidea, Kinosternoidea, Chelydridae, and Testudinoidea. Like the paracryptodira, the sinemydid-macrobaenids easily surived the K-T mass extinction, continuing to flourish in Paleogene Laurasian before giving way to more modern faunas. MAK130119
Centrocryptodira Gaffhey and Meylan, 1988: Crown Group Cryptodira and their fossil relatives
Range: Late Jurassic to Recent, Cosmopolitan
Phylogeny: Xinjiangchelyid - Crown Cryptodire clade : Chengyuchelys + Xinjiangchelyidae + Larachelus + * : Sinemydidae + Trionychoidea + Durocryptodira.
Range: Late Jurassic to Recent
Characters: Characteristics include formed (concave or convex) articulations (procoelous and opisthocoelous vertebrae) between succeeding cervical vertebrae. These well developed articulation surfaces constitute a more advanced condition then in more primitive Eucryptodires, and were made a defining feature of the Centrocryptodira. A thick floor of the canalis caroticus internus in the pterygoid is also a feature of this group. (Gaffney, 1975, Meylan 2001). This characteristic also developed independently in both the Pleurodira and Baenidae.
Comments: Gaffney co-workers' placement of the Meiolaniids is controversial, statistical-computational trees place them further towards the base of the testudine tree than other chelonians. For this reason, apart from the Gaffney school the term Centrocryptodire is no longer used. Although statistical morphological analyses support Sinemydidae as basal to extant cryptodires, molecular phylogeny instead favours a separate origin for the Trionycha, which would make the Centrocryptodira as originally defined polyphyletic MAK130119
With the increasing popularity of quantitaive methodologies and the replacement of morphology-based Hennigian cladistics and apomorphy-based clades with statistical-computational parsimony analysis and stem and crown based cladistic nomenclatural preference of restricting a familiar and morphologically well-defined taxon to the crown group only, apomorphic terms like Centrocryptodora and Polycryptodira have been rejected in favour of simply as "Cryptodira". The problem is that excessive reliance of crown clade terminology can lead to ambiguity. A well known example of this is the many definitions of Tetrapoda. In order to avoid confusion we have appended the phrase "crown group", to distinguish this taxon from the morphological (or apomorphic) Cryptodira. MAK130119
The Centrocryptodora or Crown Group Cryptodires represent a group of derived eucryptodires that are first known from the Late Jurassic of China. Of the two lineages, Yehguia tatsuensis and Sinaspiderites wimani are pantrionychids (or trionychoidea) known from Sichuan, whereas Sinemys lens is the earliest known Sinemydid , from Shandong (Rabi et al 2010 p.266). If at least some and perhaps all of the Sinemydids are durocryptodires it would solve the otherwise annoying problem of excessive ghost lineages even if it means a gread deal of convergent evolution. Chinese Jurassic eucrptodire diversity was extremely high; the different localities may imply different geographic and ecological regions.
Lourenço et al (2012), using molecular clocks calibrated with fossil taxa place the divergence between trionychoidans and non-trionychoidan cryptodires (in their paper, here Durocryptodira) at around 144.0 Mya, or in other words the Jurassic-Cretaceous boundary (Lourenço et al 2012 p.502). This would seem to be a little young as both the oldest Sinemydids and stem trionychans date to the late Jurassic. But the incongruence between the mammal fossil record and molecular clocks does not surport the premise of an invariant random mutation rate, and the same seems to be the case here.
Links: Taxon diversity
Durocryptodira: hard shelled cryptodires Danilov & Parham, 2006. Molecular, node-based definition: LCA of Testudo graeca Linnaeus, 1758, Kinosternon scorpioides (Linnaeus, 1766), Chelonia mydas (Linnaeus 1758), and Chelydra serpentina Linnaeus, 1758, but excluding Trionyx triunguis (Forskål, 1775) and Carettochelys insculpta Ramsay, 1887. From duro-, Latin for hard, and cryptodira- for cryptodires. Danilov & Parham, 2006p.577. If the molecular-phylogeny is incorrect then Durocryptodira becomes paraphyletic (or synonymous with crown group Cryptodira.
Range: fr the Early Cretaceous. Late Jurassic stem forms may have been present but had not yet evolved into extant lineages (Danilov & Parham, 2006, p.579)
Range: Fr the Late Cretaceous
Phylogeny: Centrocryptodira : Some Sinemydidae + Trionychoidea + * : Some Sinemydidae + (Kirgizemys + (Macrobaenidae + Testudinoidea + Chelonioidea-Chelydridae-Kinosternoidea clade)
Comments: Apart from Danilov and Parham, the use of the term Durocryptodira has not caught on. It is retained here only because there seems to be no other name for this molecular-sequenced clade. The absence of clear morphological traits among molecular taxa such as Durocryptodira is a well recognised feature in many higher mammalian clades derived from molecular sequencing. MAK130118
It is not unlikely that stem representatives lie among the Late Jurassic and Cretaceous Sinemydidae and Macrobaenidae, which help to fill the long stratigraphic gap betrween the early late Jurassic and the late Cretaceous. In this molecular-stratigraphic scenario, during the later middle to early late Jurassic, the testudines underwent a dramatic evolutionary radiation. Five long-lived clades emerged at this time, the Meioliinids or terrestrial horned tortoises, a primitive lineage first known from the Cretaceous, the Pleurodira or side-necked semi-aquatic forms, the Paracryptodira or unspecialised forms, the Cretaceous to Paleogene Sinemydidae-Macrobaenidae group that became the ancestors of either all extant cryptodires or of the extant hard-shelled types (Durocryptodira), and the Trionychia or advanced soft-shelled turtles, which are remarkably morphologically convergent with the durocryptodiran Kinosternoidea. Even so the position of the Trionychoidea in the phylogeny given here is extremely provisional, pending more analysis of and insights into Sinemydid and Macrobaenid phylogeny.
During the mid Cretaceous the durocryptodires seem to have undergone a sudden bursts of evolution. Lourenço et al 2012 using molecular dating, posits a rapid evolutionary radiation involving the origin of the extant Kinosternidae, Chelydridae, Chelonioidea and Testudinoidea at between 120 and 90 Mya (mid-Cretaceous), shown by the short internal branches between these clades in their phylogram. MAK130403
Horizon: Early Cretaceous of Spain
Phylogeny: Centrocryptodira : ((Kirgizemys + (Macrobaenidae + Chelonioidea-Chelydridae-Kinosternoidea Clade) + Testudinoidea)) + Trionychoidea + (Sinemydidae + *))
Comments: the western-most record of a Sinemydid-Macrobaenid grade chelonian.

Sinemydidae: Dracochelys, Manchurochelys, Ordosinemys, Sinemys
Range: Late Jurassic to Cretaceous of Asia
Phylogeny: Centrocryptodira : ((Kirgizemys + (Macrobaenidae + Chelonioidea-Chelydridae-Kinosternoidea Clade) + Testudinoidea
) + Osteopygis + Trionychoidea + (Hoyasemys + *Characters: completely formed central articulation on the cervical vertebrae.
Comments: cervical vertebrae represent a more advanced condition than Xinjiangchelys (Hirayama et al 2000 p.189). Paraphyletic as normally defined. However a smaller grouping, including Dracochelys, Manchurochelys, Ordosinemys, and Sinemys, would seem to be monophyletic, according to some statistical analyses.
In contrast to the "true" Macrobaednids, Annemys spp., Ordosemys spp., Dracochelys bicuspis, Hangaiemys hoburensis and Sinemys spp. still retain an open carotid fenestra and reduced basipterygoid process of the basisphenoid, a primitive feature homologous with that of basal turtles using topology and intermediates. (Rabi & Joyce 2012)
Graphic: Manchurochelys liaoxiensis, via Wikipedia
Range: Late Cretaceous to Early Paleocene of North America (New Jersey)
Phylogeny: Centrocryptodira : (Hoyasemys + Sinemydidae) + Trionychoidea + (Testudinoidea + (Kirgizemys + (Macrobaenidae + Chelonioidea-Chelydridae-Kinosternoidea Clade))) + *
Comments: According to Parham 2005, specimens referred to Osteopygis from Late Cretaceous to Early Paleocene marine sediments of the USA represent a chimera (parts of two species confused as the same animal), combining the head of the marine turtle Euclastes with the shell and post-crania of macrobaenids. Osteopygis emarginatus can be distinguished from Judithemys and most Asian "macrobaenids" by its larger size (shell lengths of upto 70cm), thicker shell, and the presence of plastral fontanelles. (Parham 2005 p.76). This species represents the eastern-most extent of the "macrobaenids".
Synonym: Hangaiemys (see note)
Range: Early Cretaceous of Asia
Phylogeny: Centrocryptodira : Sinemydidae + Osteopygis + Trionychoidea + (Testudinoidea + ((Macrobaenidae + Chelonioidea-Chelydridae-Kinosternoidea Clade) + *)
Comments: Basal centrocryptodiran/durocryptodiran, possibly related to panchelonoidea (Danilov & Parham, 2008), although the open open carotid fenestra (assuming synonymy of Hangaiemys and Kirgizemys) makes this a more primitive form.
[1] "I. G. Danilov, A. O. Averianov, P. P. Skutschas, A. S. Rezvyi (St. Petersburg) reported new data on the morphology of the 'macrobaenid' genus Kirgizemys known from the Early Cretaceous of Kirghizia, Buryatiya (Russia) and China. New material from the Early Cretaceous of Buryatia allowed study of the previously unknown skull of Kirgizemys. In skull and shell morphology, Kirgizemys is indistinguishable from Hangaiemys from the Early Cretaceous of Mongolia. The authors consider Hangaiemys as a junior subjective synonym of Kirgizemys." Fossil Turtle Newsletter Jan 2004
Horizon: Late Cretaceous to early Tertiary of Asia and North America
Phylogeny: Centrocryptodira : Sinemydidae + Trionychoidea + (Testudinoidea + (Kirgizemys + (Chelonioidea-Chelydridae-Kinosternoidea Clade + * : Judithemys + Macrobaena)))
Comments: As conventionally defined, the Macrobaenids are a grade of freshwater and estuarine stem cryptodires that originated in the Early Cretaceous and dies out in the late Paleocene. They consist of a range of forms more advanced than basal forms like the Baenidae and Xinjiangchelyidae, but not yet at the morphological stage of modern cryptodires, an assemblage, a grade rather than a clade, of mostly Asian, Cretaceous to early Cenozoic, taxa that would seem to be clearly related to each other, as well as to crown group cryptodira. The Macrobaenidae especially seem to be an artificial grouping of morphologically and ecologically similar types (Parham (2005)). Some recent cladistic analysis shows that the ancestors of marine turtles may be found among terrestrial forms like Sinemydids and Macrobaenids (Sterli 2008 and 2010, Pérez-García (2012)). In another tree the Sinemydids are even crownward of the chelonioidea (Slater et al (2011), p.1407). What seems to be clear is that they represent an ancestral lineage from which modern cryptrodires evolved. If the trionychoidans branched from the cryptodire stem before most other centrocryptodires, as molecular phylogeny and stratigraphy both imply, then the sinemydids and macrobaenids are included in the cryptodire crown group. And even in morphology-only analysis, some or all Sinemydids at least still seem to be crown group cryptrodira.
A narrower definition can be supposed for a smaller group such as Macrobaena and Judithemys which can be tentatively interpretred as stem chelonioid/kinosternoids and which feature a reduced carotid fenestra. This is in contrast to other Sinemydid-Macrobaenid cryptrodires which retain the more primitive or basal condition (Rabi & Joyce 2012). The closure of the carotid fenestra occurred at least three times in crown Cryptodira (the other two being trionychoids and testudinoids), although a reduced carotid fenestra is also present is primitive eucryptodires (plesiochelyids, eurysternids, and protostegids) (ibid) showing additional instances of homoplasy and phyloparallelism.
Horizon: Late Cretaceous (Campanian) Dinosaur Park Formation of Alberta, Canada
Phylogeny: Macrobaenidae : Macrobaena + *
Comments: "The morphology of Judithemys reiterates a suite of characters (large size, well-differentiated neurals reduced to eight, and greater overlap of the twelfth marginal scales onto the second suprapygal) common to Late Cretaceous-Paleocene ''macrobaenids'' and distinct from Early Cretaceous members" (Parham, J.F. & Hutchison, J.H. 2003), although its exact phylogenetic position, as for that matter that of other Asian Cretaceous-Paleogene turtles of Sinemydid/Macrobaenid grade is uncertain. It seems to be closely related to, but more advanced than, Kirgizemys
Horizon:
Phylogeny: Macrobaenidae : Judithemys + *
Comments:
Chelonioidea-Chelydridae-Kinosternoidea clade
Range: Fr the Late Cretaceous
Phylogeny: Centrocryptodira : Sinemydidae + Trionychoidea + (Testudinoidea + (Kirgizemys + (Macrobaenidae + * : Panchelonioidea + (Chelydroidea + Emarginachelys + Kinosternoidea)))
Comments: Revealed through molecular phylogeny only. Descended from ancestral Durocryptodira. Molecular phylogeny reveals a very early branching for the Trionychoidea, in which case some Sinemydids and Macrobaenids were stem durocrptodires MAK130403
| Page Back | Unit Home | Page Top | Page Next |
Page last modified MAK130403. Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners