| Palaeos | Cetartiodactyla | |
| Vertebrates | Cetacea |
| Page Back | Unit Home | Unit Dendrogram | Unit References | Taxon Index | Page Next |
| Unit Back | Vertebrates Home | Vertebrate Dendrograms | Vertebrate References | Glossary | Unit Next |
|
Abbreviated Dendrogram
LAURASIATHERIA |--FERAE `--+--PERISSODACTYLA | CETARTIODACTYLA |--Cetacea | |--Pakicetidae | `--+--Ambulocetidae | `--+--Remingtonocetidae | `--Autoceta `--Artiodactyla |--Suina | |--Hippopotamidae | `--Suoidea `--Selenodontia |--Camelidae `--Ruminantia |--Giraffoidea `--+--Bovoidea `--Cervoidea |
Contents
Overview |
These Notes contain relatively few discussions of mammals. There are a number of reasons for this, including the obvious fact that the mammals were started much later than other groups -- geologically as well as in a literary sense. However, there is at least one other reason. Mammals are extremely hard to describe since they are quite different from other, more conventional vertebrates, and have acquired some absurdly complex specializations.
No group illustrates the truth of this proposition (or at least the utility of this excuse) better than the Cetacea. The Cetacea are, of course, best known from their modern representatives, the porpoises, dolphins and whales. Their specific diversity is not particularly impressive. About 80 species survive. There are more species of cichlid fishes in any reasonably large African lake than there are cetaceans in the entire world. However, most members of the Cetacea share a suite of adaptations that are so unique and complex that we have great difficulty in understanding what they are, much less how they work.
This means that, from the point of view of an evolutionary biologist, cetaceans are very difficult to discuss. Evolutionary biology works in a complex set of dimensions which we may refer to as phylospace. A point in phylospace defined by taxon, time and space. By ever-better understanding of the points we try to understand the flow of life from Archaean blobs to African elephants. But each of the dimensions we use to define the points (a) represents a range, rather than a point and (b) is a gestalt of fractional dimensions.
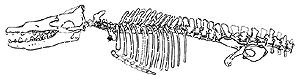 For
example, consider the Protocetidae.
This is a group of about 8 early cetacean species, including Rodhocetus
shown in the figures. The protocetids are centered on the middle Eocene and
distributed around the southern and western margins of the Tethys Sea: roughly
from Pakistan to West Africa. Even at this level, we run into significant
problems, since the group may not be monophyletic, some parts seem to stretch
into the Lower and Upper Eocene, and there is a disputed report of a protocetid
in North America. The phylodimension of time is a function of things like
geological stratigraphy and correlation, depositional l anomalies, taphonomic
biases, post-depositional transport, and so on. The phylodimension of space is
particularly important in this group, since it suggests that the protocetids
were the first cetaceans to break out of the eastern Tethys homeland. This, in
turn, suggests that they were fully at home in an open marine environment, if
not quite ready for prime time in the open Atlantic. However, this is a delicate
chain of inference. We say that protocetids were distributed from Pakistan to
Nigeria, but we actually know only that protocetids are found at several
locations in the Indian subcontinent, abundantly at one location in Egypt, and
that there is a published report of findings in Nigeria. While unlikely, it
remains possible that these are the only locations with protocetids --
that they remained coast-huggers with only a few centers of population at
specific favorable locations.
For
example, consider the Protocetidae.
This is a group of about 8 early cetacean species, including Rodhocetus
shown in the figures. The protocetids are centered on the middle Eocene and
distributed around the southern and western margins of the Tethys Sea: roughly
from Pakistan to West Africa. Even at this level, we run into significant
problems, since the group may not be monophyletic, some parts seem to stretch
into the Lower and Upper Eocene, and there is a disputed report of a protocetid
in North America. The phylodimension of time is a function of things like
geological stratigraphy and correlation, depositional l anomalies, taphonomic
biases, post-depositional transport, and so on. The phylodimension of space is
particularly important in this group, since it suggests that the protocetids
were the first cetaceans to break out of the eastern Tethys homeland. This, in
turn, suggests that they were fully at home in an open marine environment, if
not quite ready for prime time in the open Atlantic. However, this is a delicate
chain of inference. We say that protocetids were distributed from Pakistan to
Nigeria, but we actually know only that protocetids are found at several
locations in the Indian subcontinent, abundantly at one location in Egypt, and
that there is a published report of findings in Nigeria. While unlikely, it
remains possible that these are the only locations with protocetids --
that they remained coast-huggers with only a few centers of population at
specific favorable locations.
 The
most difficult and most fractured dimension, however, is the taxon itself. Using
a cladistic approach, as in these Notes, each taxon is characterized by a myriad
of character states. Cladistics has allowed us to deal with taxa more
objectively, but the evolution of complex, soft tissue organ systems is
difficult to trace from isolated osteological characters. [2].
And this is so even when we understand how the systems work in living members of
the same group. In whales, our understanding is still very incomplete. Consider
one of the more conspicuous characteristics of cetaceans, which also turns out
to be one the largest questions in cetacean biology: echolocation. As we will
see, this is not just a matter of sensory physiology, but has very deep
implications for what it means to be a cetacean.
The
most difficult and most fractured dimension, however, is the taxon itself. Using
a cladistic approach, as in these Notes, each taxon is characterized by a myriad
of character states. Cladistics has allowed us to deal with taxa more
objectively, but the evolution of complex, soft tissue organ systems is
difficult to trace from isolated osteological characters. [2].
And this is so even when we understand how the systems work in living members of
the same group. In whales, our understanding is still very incomplete. Consider
one of the more conspicuous characteristics of cetaceans, which also turns out
to be one the largest questions in cetacean biology: echolocation. As we will
see, this is not just a matter of sensory physiology, but has very deep
implications for what it means to be a cetacean.
Echolocation is best understood in the dolphin, Tursiops, for the
simple reason that dolphins (unlike many cetaceans) do not require something
like a water-filled version of Barcelona's Nou
Camp Stadium in which to conduct experiments. 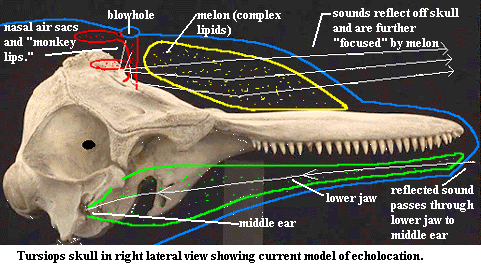 The
ability of dolphins to identify the location, composition and three dimensional
structure of small objects at considerable distances is nearly beyond belief. It
has been learned only recently that dolphins produce the "clicks" and
"whistles" of echolocation -- not in the larynx -- but probably in
nasal air sacs and associated valves ("monkey lips") opening on the
passage to the blowhole. According to current thinking, the sounds are then
reflected from, and focused by, the curved surface of the skull, and then
further focused by passing through the complex fats of the "melon"
which creates the familiar bulging "forehead" of dolphins. The result
is a sort of highly directional flashlight beam of sound. In addition, the melon
helps transduce the airborne vibrations in the air sacs into water-borne
vibrations. [3] Returning, reflected sound is
picked up by the lower jaw and conducted to the middle ear, which is
acoustically insulated from the outgoing sounds.
The
ability of dolphins to identify the location, composition and three dimensional
structure of small objects at considerable distances is nearly beyond belief. It
has been learned only recently that dolphins produce the "clicks" and
"whistles" of echolocation -- not in the larynx -- but probably in
nasal air sacs and associated valves ("monkey lips") opening on the
passage to the blowhole. According to current thinking, the sounds are then
reflected from, and focused by, the curved surface of the skull, and then
further focused by passing through the complex fats of the "melon"
which creates the familiar bulging "forehead" of dolphins. The result
is a sort of highly directional flashlight beam of sound. In addition, the melon
helps transduce the airborne vibrations in the air sacs into water-borne
vibrations. [3] Returning, reflected sound is
picked up by the lower jaw and conducted to the middle ear, which is
acoustically insulated from the outgoing sounds.
This scheme leaves a good many unanswered questions. Why does the skull reflect sound? Bone is a notoriously good conductor of sound. How is sound focused by the melon? Why isn't the focusing affected by temperature? Why isn't the information in the faint, reflected sound signal hopelessly hashed by funneling through the lower jaw? How can the brain decode this extremely complicated information into a three dimensional picture in real time? Dolphins are not only able to manage this trick, but may also be able to echolocate with random, ambient sounds. Finally, given that the echoes are equally available to other members of these gregarious species, what are the social implications in these intelligent creatures of a perceptive sense (a) that amounts to partial telepathy (not to mention X-ray vision) and (b) permits the group to build a single "view" of the surroundings by combining individual perceptions, where (c) the medium used overlaps with sound communications?
 How
does one study the evolution of such a phenomenon? The known facts are, even in
these days of virtually unlimited computing power, absolutely awe-inspiring. We
barely understand how it works and lack the neurological capacity to ever
appreciate the sense impression itself, much less the social, cognitive, and
even artistic implications of having such a shared sense. So, we can make
guesses about the anatomy, social life and sensory ability of protocetids; but
we literally lack the brains, as well as the data, necessary to appreciate how
the development of this sort of biological factor affected their mode of life.
How
does one study the evolution of such a phenomenon? The known facts are, even in
these days of virtually unlimited computing power, absolutely awe-inspiring. We
barely understand how it works and lack the neurological capacity to ever
appreciate the sense impression itself, much less the social, cognitive, and
even artistic implications of having such a shared sense. So, we can make
guesses about the anatomy, social life and sensory ability of protocetids; but
we literally lack the brains, as well as the data, necessary to appreciate how
the development of this sort of biological factor affected their mode of life.
Not that this will stop anyone from trying, of course. Nor should it. But the job is certainly more daunting than, say, working out a sequence of cranial bone homologies in reptiles; and mammalian evolution seems to be full of impossible questions of this kind. The remarkable thing is that we have made much progress at all. --ATW 010515
Notes: [1] As Thewissen et al. (2001) note, either whales are the sister group of artiodactyls and independently evolved mesonychid-like dentition, or they are the sister of mesonychids and independently evolved the artiodactyl ankle.
[2] For an outstanding example of a successful attack on this kind of problem, see Hutchison (2001) and Hutchison (2001a).
[3] This is precisely the reverse of the impedance-matching problem discussed in connection with the middle ear.
[4] This essay would seem to owe a lot to Zimmer (1998). A recent (030120) re-reading of this source has convinced us that it would be appropriate to acknowledge that debt more explicitly.
References: Hutchinson (2001); Hutchinson (2001a); Nowak (1991); Thewissen et al. (2001); Vaughan et al. (2000); Zimmer (1998). 011027
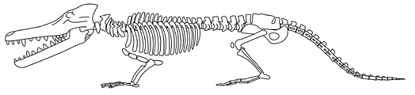 Cetacea:
Ambulocetus, Himalayacetus.
Cetacea:
Ambulocetus, Himalayacetus.
Range: from the Early Eocene [BG98]. These authors estimate the divergence from Artiodactyla to be latest Cretaceous or Early Paleocene, with adaptation to a primarily aquatic lifestyle by the beginning of the Eocene.
Phylogeny: Cetartiodactyla : Artiodactyla + * : Pakicetidae + (Ambulocetidae + (Remingtonocetidae + Protocetidae)).
Characters: Body primitively under 3m, now 1.2 - 30m and 20kg to 160T; tear-drop body shape; elongate skull; zygomatic arch turns ventrally at origin on maxilla; nares usually at highest point of skull; high sagittal crest and marked lambdoid crest; occipital articulation simple or may be fused; braincase narrow; tympanics (bulla), petrosals & auditory ossicles pachyostotic [T98]; ventral floor of external auditory meatus absent [L98]; tubular portion of ectotympanic contribution to external auditory meatus reduced [L98]; bulla formed from ectotympanic [C88]; involucrum present [$L98]; bulla posterior process (derived from posterior crus of epitympanic ring) tapering posteriorly as part of external auditory meatus [L98]; bulla with sigmoid process [T98]; primitively, sigmoid process is simple plate, articulating with basicranium on the squamosal [L98]; bulla with lateral furrow [$L98]; massive auditory bulla formed from ectotympanic, primitively attached to squamosal, basioccipital and paroccipital; after initial expansion, size of bulla actually decreases as a % of body mass [G98]; middle ear "rotated" [T98]; incus "twisted" [Z98]; incus mallear articulation faces anteriorly, rather than anteromedially [TH93]; incus pachyostotic, with crus longum very short [TH93]; and fossa for malleus separate from epitympanic recess [T98]; groove for tensor tympani between promontorium and the tegmen tympani [T98]; reduced "occlusal basins" (talonid?) and enlarged shearing surfaces [T98]; premolars thin & blade-like; P4 protocone absent [T98]; upper molars retain 3 roots; molars with large medial hypocone; vascularized region between medial portions of upper molars; may have dorsal fin; horizontal tail flukes; forelimbs generally modified to form flippers for steering; pelvic girdle, where present, used only for external reproductive organs; hindlimbs reduced or absent; sweat and sebaceous glands absent; fibrous layer with fat and oil (blubber) beneath skin; bones are spongy & oil-filled; echolocation and sound communication common; locomotion by horizontal undulation using highly developed axial muscle system (faster than actinopterygians); complex thermoregulatory adaptations; direct connection between nares & lungs; most taxa gregarious; modern groups with high intelligence, some probably self-aware; single births typical; extended parental care.
Image: Ambulocetus from Dr. Hans Thewissen's Ambulocetidae.
Links: Cetaceans; Time of origin of the Cetacea; Systematic Biology 48(1) Abstracts; Eocene Cetaceans; ambulo.htm; Pb234The.htm; Ambulocetidae Research; Whale Evolution- Enchanted Learning Software; Order Cetacea (ADW); Order Cetacea; cetacea (French); Whales, Dolphins, and Porpoises of the Cetacea Order (very basic); CETACEA- Whales, Dolphins, and Porpoises (Walker's extract); Cétacés (baleines, dauphins et marsouins) (French); Cetacea - Wikipedia; Order CETACEA (skull images); Douglas J. Futuyma, "Cetacea Evolution," Evolutionary Biology, ...; Cetacea.
References: Bajpai & Gingerich (1998) [BG98]; Carroll (1988) [C88]; Gingerich (1998) [G98]; Luo (1998) [L98]; Thewissen (1998) [T98]; Thewissen & Hussain (1993) [TH93]; Zimmer (1998) [Z98]. ATW031016.
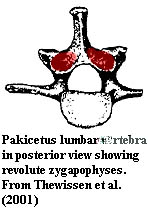
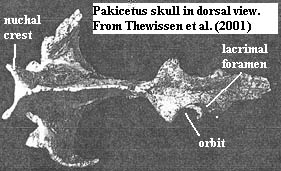 Pakicetidae:
Ichthyolestes, Nalacetus, Pakicetus
Pakicetidae:
Ichthyolestes, Nalacetus, Pakicetus
Range: Early Eocene to Middle Eocene of South Asia.
Phylogeny: Cetacea : (Ambulocetidae + (Remingtonocetidae + Protocetidae)) + *.
Characters: up to medium dog size [T98]; nares at tip of snout
[T+01] or over incisors [T98]; lacrimal foramen present [T+01]; $
orbits closely spaced and "frontated," i.e. face
dorsally, but are not at most dorsal point of skull [T+01] [T98]; hypoglossal
foramen separate from jugular foramen [T98]; external auditory
meatus opens low on skull as in land animals [T+01]; dense tympanic bulla
[GU98]; bulla still attached to squamosal, basioccipital & paroccipital
process [C88]; bulla posterior process thin vertical plate forming posterior
wall of external auditory meatus [L98]; petrosal was firmly integrated in the basicranium (poor directional
hearing in water) [G98]; probably flat tympanic membrane as
in terrestrial animals [T+01]; middle ear with involucrum
and plate-like sigmoid process [T+01]; sigmoid process plate-like, continuous
with (& forming) anterior wall of external auditory meatus [L98]; ear ossicles dense and enlarged
(underwater or earthborne vibration) [T+01]; incus intermediate in morphology
between modern artiodactyls and cetaceans (suggests poor  hearing in water)
[GU98]; palatine fissures (??) present [T98]; mandibular foramen small (not large and adapted for underwater hearing)
[GU98] [T98] [T+01]; premolars serrated triangular blades, like shark teeth [C88]; teeth retaining complex morphology and functional occlusion, with
larger protocones but otherwise similar to protocetids
and basilosaurids [GU98]; upper molars
triple-rooted [C88]; lower molars with distinct paraconid & metaconid [T98]; lower
molars with distinct protoconid & hypoconid [C88]; hypoconid is only cusp on
talonid [T98]; dental wear consisting of vertical gouges
[T+01]; relatively long cervical vertebrae [T+01]; lumbar
and caudal vertebrae of moderate length [T+01]; lumbar spine with revolute
zygapophyses (presumably permitting dorsoventral flexion only) [T+01]; four
strongly fused sacral vertebrae [T+01]; $ large supraspinous
fossa and small acromion process [T+01]; humerus long & slender [T+01];
deltopectoral crest minimal [T+01]; wide, tightly constrained hinge joint for
radius & ulna [T+01]; olecranon small (<12% of ulna length) [T+01];
innominate large, with ischial portion longer than ilium [T+01]; tibia long,
with short tibial crest [T+01]; large tibial trochlea of astragalus forms tight
hinge joint rotating in dorso-plantar plane [T+01]; "the sustentacular
facet is a hinge that also rotates dorso-plantarly" [T+01] (??); ectal
facet small and lateral [T+01]; long calcaneal tuber (cursorial) [T+01]; narrow,
oblique (?) cuboid facet of calcaneum (cursorial) [T+01]; slender metatarsals
[T+01]; found in river & estuarine areas in association with land animals
[GU98].
hearing in water)
[GU98]; palatine fissures (??) present [T98]; mandibular foramen small (not large and adapted for underwater hearing)
[GU98] [T98] [T+01]; premolars serrated triangular blades, like shark teeth [C88]; teeth retaining complex morphology and functional occlusion, with
larger protocones but otherwise similar to protocetids
and basilosaurids [GU98]; upper molars
triple-rooted [C88]; lower molars with distinct paraconid & metaconid [T98]; lower
molars with distinct protoconid & hypoconid [C88]; hypoconid is only cusp on
talonid [T98]; dental wear consisting of vertical gouges
[T+01]; relatively long cervical vertebrae [T+01]; lumbar
and caudal vertebrae of moderate length [T+01]; lumbar spine with revolute
zygapophyses (presumably permitting dorsoventral flexion only) [T+01]; four
strongly fused sacral vertebrae [T+01]; $ large supraspinous
fossa and small acromion process [T+01]; humerus long & slender [T+01];
deltopectoral crest minimal [T+01]; wide, tightly constrained hinge joint for
radius & ulna [T+01]; olecranon small (<12% of ulna length) [T+01];
innominate large, with ischial portion longer than ilium [T+01]; tibia long,
with short tibial crest [T+01]; large tibial trochlea of astragalus forms tight
hinge joint rotating in dorso-plantar plane [T+01]; "the sustentacular
facet is a hinge that also rotates dorso-plantarly" [T+01] (??); ectal
facet small and lateral [T+01]; long calcaneal tuber (cursorial) [T+01]; narrow,
oblique (?) cuboid facet of calcaneum (cursorial) [T+01]; slender metatarsals
[T+01]; found in river & estuarine areas in association with land animals
[GU98].
Image: Pakicetus life reconstruction by Carl Buell, and adapted from http://www.neoucom.edu/Depts/Anat/Pakicetid.html.
Links: Pakicetidae (Thewissen's web page and Best on the Web); The Emergence of Whales, Chp. 1; Likelihood Estimation of the Time of Origin of Cetacea and the Time of Divergence of Cetacea and Artiodactyla; Archaeocetes- Archaic Whales, Dolphins and Porpoises; Élet és tudomány (Hungarian); Cetacea.de | Palaeocetologie Indopakistans (German); MAMMALIAN EVOLUTION MEETING; Chapter 19 Cenozoic Mammals- Guilds and Trends; A new Eocene archaeocete (Mammalia, Cetacea) from India and the ...; Limbs in whales and limblessness in other vertebrates- mechanisms ...; Science -- Gingerich et al. 293 (5538)- 2239.
References: Carroll (1988) [C88]; Gingerich & Uhen (1998) [GU98], Luo (1998) [L98]; Thewissen (1998) [T98]; Thewissen et al. (2001) [T+01]. ATW040720.
Ambulocetidae: Ambulocetus, Gandakasia
Range: Middle Eocene of South Asia.
Phylogeny: Cetacea :: (Remingtonocetidae + Protocetidae) + *.
Characters: Ambulocetus mass variously estimated at 140-720 kg [G98]; supraorbital region concave & supraorbital shield is absent [T98]; orbits dorsally placed, not merely frontated [T+01] [U98] (contra? orbits face laterally and are close to sagittal plane [$T98]); $ pterygoid process of sphenoid approximately same height as braincase [T98]; mandibular foramen enlarged (presumably for underwater hearing) to half height of mandible [T+01]; upper molars with large parastyle [T98]; metaconid absent [T98]; flexible, relatively unconstrained vertebral column [T+01]; cervical vertebrae not longitudinally compressed [U98]; functional torso limited to lumbars [B98]; fewer lumbar than thoracic vertebrae [B98]; caudal vertebrae do not show abrupt shortening of more derived whales [U98]; no caudal peduncle or fluke [B98]; large olecranon process [T+01]; radius & ulna robust & cylindrical to oval in cross-section [U98]; radius immovable relative to ulna [U98]; large, curved articular surfaces on carpals, permitting significant flexibility [U98]; magnum & trapezoid unfused [U98]; pelvis robust [B98]; hind limbs very large [U98]; feet much more robust than pakicetids [T+01]; feet, not tail, used as hydrofoils [T98]; may have been fresh water or partially fresh water organisms [T98]; locomotion by flexion & extension of lumbar region & using large feet to provide propulsion [B98].
References: Buchholtz (1998) [B98]; Gingerich (1998) [G98]; Thewissen (1998) [T98]; Thewissen et al. (2001) [T+01]; Uhen (1998) [U98]. ATW011027.
 Remingtonocetidae:
Andrewsiphius, Attockicetus, Dalanistes, Kutchicetus, Remingtonocetus.
Remingtonocetidae:
Andrewsiphius, Attockicetus, Dalanistes, Kutchicetus, Remingtonocetus.
Range: Middle Eocene (Lutetian) of India & Pakistan [FB94].
Phylogeny: Cetacea ::: Protocetidae + *.
Characters: skull very long [U98]; rostrum very long and thin [BT98] [FB94] [G98] [$T98]; nares somewhat retracted [G98] but only slightly [BT98]; supraorbital shield present (?!) [T98]; orbits small [BT98] [$T98]; posterior skull wide (but not in Andrewsiphius [BT98]); ears displaced laterally & well separated [$T98]; braincase low & wide [BT98]; falcate process of basioccipital variably present (very strong in Remingtonocetus and absent in Andrewsiphius) [BT98]; oblong tympanics [$T98]; expansion of tympanic sinuses controversial & perhaps absent [FB94] (contra [BT98]? [B98]?); convex posterior palate extending ventrally beyond tooth crowns [BT98] [$T98]; mandibular symphysis fused [U98]; mandibular symphysis very extensive [$T98]; mandibular foramen enlarged (presumably for underwater hearing) [T98]; cheek teeth placed far forward of orbits [FB94]; protocone small [$T98]; upper molars with large parastyle [T98]; metaconid absent [T98]; flexible, relatively unconstrained vertebral column [T+01] (contra [G98]); vertebral formula 7+14+5+4 (caudals unknown) [B98]; cervical vertebrae not longitudinally compressed [U98]; cervicals at least as long as tall [B98]; thoracics increase gradually in length & are significantly wider than high (ratio 1.4-1.7) [B98]; thoracic vertebrae with long neural spines (forelimbs potentially weight-bearing) [G98]; lumbar and sacrals of similar length [B98]; 4 sacral vertebrae strongly fused and attached to hip [G98] [U98]; long caudal and lumbar vertebrae (moved by body undulation?) [T+01]; caudals broader than Ambulocetus [B98]; peduncle and fluke "almost certainly" absent [B98]; pelvis large [G98]; femur robust [G98] [T98]; hind limb robust [B98]; aquatic locomotion by flexing lumbars & foot propulsion as in Ambulocetus, with secondary role for tail [B98]; able to move on land [B98]; ambush predator? [T98].
Note: May be first marine cetacean. Thewissen [T+01] and Gingerich [G98] speculate that remingtonocetids may have specialized as ambush predators of marine shorelines. These are really weird whales. To us, they look more like longirostrine piscivores.
Links: Remingtonocetid.
Image: cranium of Remingtonocetus from JGM Thewissen. Apparently adapted from [BT98].
References: Bajpai & Thewissen (1998) [BT98]; Buchholtz (1998) [B98]; Gingerich (1998) [G98]; Fordyce & Barnes (1994) [FB94]; Thewissen (1998) [T98]; Thewissen et al. (2001) [T+01]; Uhen (1998) [U98]. ATW010512.
| Page Back | Unit Home | Page Top | Page Next |
checked ATW051030