Cambrian Epoch 2
| Palaeos: Paleozoic |
Terreneuvian Epoch & Cambrian Epoch 2 |
|
| Cambrian Period | "Early Cambrian": 1 |
| Page Back | Back: Ediacaran | Up: Cambrian | Unit Home | |
| Page Next | Next: Middle Cambrian | Next: Ordovician | Down: Fortunian | Timescale |
This was a time of great change across a wide spectrum, from tectonic (the breakup of the early Rodinia and Pannotia supercontinents) through geographical (marine transgressions as the ocean invaded the land creating shallow continental seas that allowed primitive life to flourish), atmospheric, (rising oxygen and falling carbon dioxide levels), biogeochemical (changing ocean chemistry, promoting biomineralization; especially the upwelling of high phosphorous levels (large phosphorite deposits), enabling organisms to develop skeletons of calcium phosphate), and ecological (the appearance of predation and algae grazers, which meant an end of the Ediacaran idyll). This combination of events would seem to have been responsible for the sudden and remarkable and diversification of metazoan life at this time.
The Early Cambrian was a time of incredible diversification. Quite possibly, this process had begun earlier, but the fossil record first reveals the results during this relatively brief interval of 30 My or so. It seems to have proceeded in three stages, sometimes referred to by the names of three Siberian Cambrian stages: the Manykian, Tommotian and Atdabanian. At the beginning of the epoch, we have simple worm traces, followed shortly by tiny, rather simple shells, the so-called "Small Shelly Fauna" of the earliest Cambrian, or Manykian Age. This was followed by a much more diverse group of shelly animals often referred to as the Tommotian fauna. The relationships of Tommotian animals to living groups are not understood, but it is likely that mollusks also evolved at this time. The Tommotian animals disappeared rather quickly and were followed by the "Cambrian Explosion" of modern groups in the last half of the Early Cambrian (Atdabanian Age). By the end of the Early Cambrian, almost all of the modern phyla had evolved: mollusks, annelid worms, arthropods (including the trilobites), echinoderms, and chordates, to name but a few.
Thus, in the space of about 17 My, roughly from 530 to 513 Mya, the pattern was set for almost all subsequent macroscopic animal life. Many writers have marveled at the amount of new diversity. Stephen Gould’s popular book on the Burgess Shale, of the Canadian Middle Cambrian, Wonderful Life, celebrates this explosion in some detail. Gould (1989). What we are now learning is even more startling -- the extraordinary durability of this new diversity. Since this was the first radiation of metazoans, its not really surprising that this new life form, expanding into an ecological vacuum, should produce many new varieties. What is a bit harder to explain is why so many of these animals reflect groups which survived for hundreds of millions of years. Most explosive radiations seem to generate a lot more in the way of unsuccessful early models. Perhaps these forms were weeded out in the Manykian and Tommotion Ages. Then again, perhaps we just haven’t found enough fossils yet. The remarkable fossils of Chenjiang, of latest Early Cambrian age, are just now being described in detail. These fossils may eventually give us an entirely new perspective on this key period of early metazoan life.
We do know that the seas rose continuously throughout the Early Cambrian. Higher water is almost always good for life, and this was particularly true during the Paleozoic. Here’s why. The deep oceans are not very useful for life. Plants and photosynthetic bacteria form the base of the foodchain everywhere. Only the top 100 meters or so of the deep oceans are reached by light and can support these organisms. In addition, the deep oceans provide no substrate for plants to attach and get big. However, when the seas rise, they spill over the broad flat coastal plains of the continental shelves. Thus, a small rise in sea level means a lot more area covered by water. This new area of continental sea is shallow -- almost all of it within the "photic zone" in which plants can live. These sunlit continental seas also provide a substrate on which photosynthesizers can attach and grow large. Thus even relatively small increases in sea level can have huge effects on the productivity of the oceans.
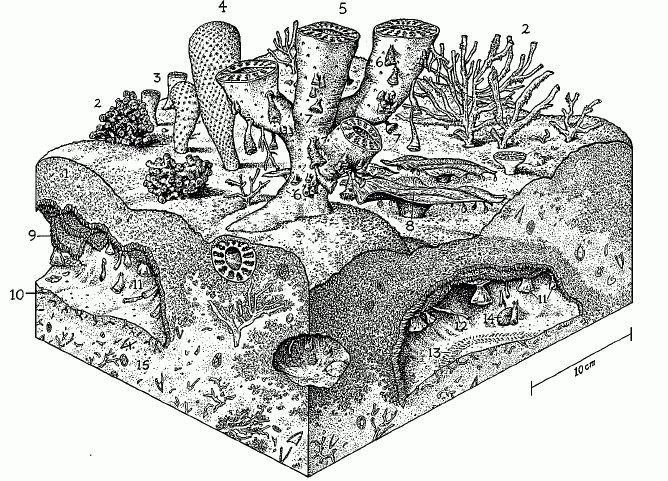
Shallow seas also provide a substrate for reef-builders. We tend to think of coral or oyster reefs as picturesque, ecological reservoirs of diversity confined to a relatively small band of tropical waters. But we live in a very unusual corner of Earth history. By comparison with most of the Phanerozoic, our age is bitterly cold and dry. In many, more typical portions of planetary space-time, the shallow seas were almost all warm and dominated by vast reefs which formed complex three-dimensional marine arcologies in wide belts extending for of hundreds, or even thousands of kilometers along the ancient coasts. The Early Cambrian saw the first large reef complexes of this type, although they would be unimpressive by Mesozoic standards. The reef-builders of that epoch were the lacy, vase-shaped archaeocyaths, probably related to sponges, along with other organisms whose affinities are not yet understood.
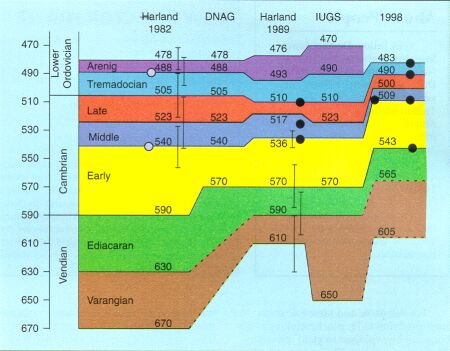
Around 1990 it became evident that most of the published radiometric data on the Cambrian were based on Rubidium-Strontium, which gave ages that
were up to 50 million years too high. This created a series of drastic revisions, in the age assignment of the Cambrian in general, and especially its lower boundary as
shown in the above figure (for more detailed information see ![]() Cambrian Stratigraphy) Overall the presumed length of the Cambrian has been reduced from almost
100 million years to little more than half of that. However, it can be seen that the Early Cambrian (yellow band in this diagram) takes the
lion's share and in fact remains a longer interval than both the Middle and Late Cambrian combined.
Cambrian Stratigraphy) Overall the presumed length of the Cambrian has been reduced from almost
100 million years to little more than half of that. However, it can be seen that the Early Cambrian (yellow band in this diagram) takes the
lion's share and in fact remains a longer interval than both the Middle and Late Cambrian combined.
The Cambrian of the Siberian Platform and adjacent areas are among the best studied Cambrian sequences, and the traditional Siberian Early Cambrian stages (Tommotian, Atdabanian, Botomian, Toyonian) are often used for global comparisons although no formal decision has been made. In in 1992 the Nemakit-Daldynian, now called the Fortunian Stage, was added for the most basal strata, preceeding the Tommotian. The entire series is shown in the table here.
During the first 10 million years of the Cambrian Period the Tommotian fauna was replaced by a more diverse biota of larger metazoans participating in more complex communities. The Cambrian fauna was dominated by low-level suspension feeders such as the nonarticulate brachiopods and eocrinoids together with monoplacophoran and hyolith mollusks. Two parazoan groups developed colonial strategies: the archaeocyaths and possibly the sponges. Colonies tend to develop iteratively with new iterative units or modules derived by continuous growth from existing units. The colonial or iterative body plan thus contrasts with the unitary or solitary life mode of most organisms. A number of groups show clear trends towards a greater integration of individuals within the colony and in some cases a differentiation of functions across the colony.
MAK020516.
 Notes from the literature:
Notes from the literature:
[S]tudies of Cambrian fossiliferous strata in Siberia, China, Europe, and Australia have yielded an unexpected range of well-preserved phosphatic microfossils. Most are either tiny shells or the disarticulated components (sclerites) of a protective armour comprised of many parts (Bengtson et al., 1990). These microfossils have become familiar as the "small shelly fossils." Most are original phosphatic skeletons or phosphatic copies of other kinds of hard parts ... .
Stimulated in part by an international endeavour to define the Precambrian-Cambrian boundary for the global geological time scale, studies of Cambrian fossiliferous strata in Siberia, China, Europe, and Australia have yielded an unexpected range of well-preserved phosphatic microfossils. Most are either tiny shells or the disarticulated components (sclerites) of a protective armour comprised of many parts (Bengtson et al., 1990). These microfossils have become familiar as the "small shelly fossils." Most are original phosphatic skeletons or phosphatic copies of other kinds of hard parts ... .
Grotzinger et al. (1998) (abstract):
Abundant calcified fossils occur in the terminal Proterozoic Nama Group, Namibia. The fossils are constrained to occur stratigraphically below the first occurrence of Phycodes pedum, overlap in range with known Ediacaran fossil impressions, and extend between ash beds dated at 549 and 543 Ma. At least six distinct morphotypes have been identified to date, suggesting that the proportional representation of calcareous taxa in the terminal Proterozoic fauna was not markedly different from that observed in younger periods. New taxa include branching forms up to 1 cm in diameter that display hexagonal symmetry. This symmetry recallsthe tri-radiate geometry of several prominent terminal Proterozoic and basal Cambrian taxa, including Tribrachidium and Anabarites. Other forms include open cups possibly representative of simple asconoid (vase-shaped) sponges; simple tubes that are functionally (if not necessarily phylogenetically) comparable to sabellid worm tubes, possibly indicating the presence of anterior tentacles or lophophore-like appendages; and more complex multichambered forms. In terms of abundance, the simple cups and tubes are most numerous, followed by the complex branching and multichambered forms, and then the previously described Cloudina. All calcified fossils in the Nama appear to represent benthic organisms that lived in close association with thrombolitic and stromatolitic reefs and biostromes, suggesting a strong substrate preference. Generally, they are rare or absent in other facies. Although the forms with hexagonal symmetry may be related to Anabarites, most Nama calcareous fossils disappear near the PC-C boundary and do not appear to be closely related to Cambrian small shelly fossils. For the most part, they are as enigmatic as the canonical Ediacaran biota.
 Haas (1981)
paraphrase:
Haas (1981)
paraphrase:
Small shelly fossils near the base of the Cambrian mark the transition to a skeletonized fauna and the metazoan-dominated Phanerozoic fossil record. The recovery of articulated specimens composed of multiple sclerites discussed above, such as Wiwaxia, Halkieria and Microdictyon, suggests that much of the remaining "small shelly fauna" represent elements similarly employed as ectodermal armor in bilaterian Metazoa that have yet to be recovered in an articulated form. In addition, recent finds of cap-shaped shells composed of fused spicules of the early Cambrian age (Bengtson, 1992) support an argument of fusion of individual skeletal elements to form sclerites, plates, or shells.
Thus, the engrailed data reported here, in combination with previous scenarios for the formation of ectodermal armor and recent fossil discoveries, suggest a singular evolution of invertebrate skeletons near the base of the Cambrian, followed by subsequent loss in several lophotrochozoan and ecdysozoan lineages. ... Such an interpretation, if substantiated, would have important implications for the Cambrian radiation, as it would constrain readily fossilizable exoskeletons to a single lineage, leading to a monophyletic clade of bilaterians. This would lead to a closer association of the bilaterian diversification with the base of the Cambrian. ...
Onychophorans are thought to be the sister taxon of arthropods and are segmented. However, onychophorans lack engrailed expression in their dermis. Instead, expression is observed in the posterior half of the developing limb and in a segmental pattern in the lateral mesoderm. The limb staining suggests shared ancestry of the onychophoran and arthropod limbs. However, given the close relationship of Arthropoda and Onychophora, and their segmented body plans, the lack of segmental ectodermal expression in Onychophora suggests that the ancestral role of engrailed was not segmentation; this absence may be a consequence of evolutionary loss of skeletons. Onychophoran dermis lacks a chitinous cuticle; thus Onychophora lack an exoskeleton. Furthermore, Cambrian fossils thought to be stem group onychophorans, such as Microdictyon, Hallucinogenia, and Xenusion, bear skeletal elements above the limb on each segment. Therefore, the absence of engrailed transcription in the ectoderm of modern Onychophora could well be a consequence of evolutionary loss of exoskeletal elements coincident with the loss of a component of engrailed expression associated with skeletal boundaries. In addition, the Cambrian Palaeoscolex, an ecdysozoan (Conway Morris 1997), bears more extensive ectodermal sclerites than modern pripulids, and Wiwaxia and Halkieria (Conway Morris & Peel 1995) bear more armor than many modern lophotrochozoa. These observations are consistent with the loss of skeletal elements in a number of lineages since the early Cambrian Period.
©Chris Clowes, 2002. CC020114, ATW070120
| Page Back | Unit Home | Page Up | Page Top | Page Next |