Porifera - 2
 Sponges, Fungi, and Ediacarans: the Neoproterozoic "Fauna." There is now relatively general agreement that the Porifera are paraphyletic. That is, the last common ancestor of all sponges was also one of our own ancestors. As Sperling et al. (2006) have pointed out, sponge paraphyly implies that many of the features we thought were sponge specializations must in fact be primitive for all metazoans. In particular, the water-canal system and the poriferan sessile, benthic mode of life are probably primitive for Metazoa. Personally, we were considerably relieved to learn that our recurrent desire to sit motionless in a liquid medium, indiscriminately absorbing ambient hors-d'oeuvres, is thus not necessarily a consequence of advanced degeneracy, but merely a plesiomorphic desire inherited from our sponge ancestors.
Sponges, Fungi, and Ediacarans: the Neoproterozoic "Fauna." There is now relatively general agreement that the Porifera are paraphyletic. That is, the last common ancestor of all sponges was also one of our own ancestors. As Sperling et al. (2006) have pointed out, sponge paraphyly implies that many of the features we thought were sponge specializations must in fact be primitive for all metazoans. In particular, the water-canal system and the poriferan sessile, benthic mode of life are probably primitive for Metazoa. Personally, we were considerably relieved to learn that our recurrent desire to sit motionless in a liquid medium, indiscriminately absorbing ambient hors-d'oeuvres, is thus not necessarily a consequence of advanced degeneracy, but merely a plesiomorphic desire inherited from our sponge ancestors.
Actually, this description may not fit the Porifera so much as their Ediacaran competition. The poriferan ability to circulate water actively and suck out its carbon content efficiently may well have been a key reason why we are descended from sponges, rather than from "vendobiota." Sperling et al. (2006). But this comparatively energetic activity may have been a later development. The Ediacaran fauna probably represent a number of different lineages which all diverged from the Eukarya at about the same time, and from very close to the same place in phylospace, as the Metazoa and Fungi. This is the most reasonable explanation for the profound observation of Peterson et al. 2003) that Ediacarans are often remarkably fungal. Perhaps, originally, sponges were simply  another one of these lineages. Hexactinellid sponges, in particular, have at least two important characteristics which are unique among animals, but (in our view) are shared with Fungi -- secondarily syncytial tissues and "plugged pore" junctions. Leys et al. 2006). [13].
another one of these lineages. Hexactinellid sponges, in particular, have at least two important characteristics which are unique among animals, but (in our view) are shared with Fungi -- secondarily syncytial tissues and "plugged pore" junctions. Leys et al. 2006). [13].
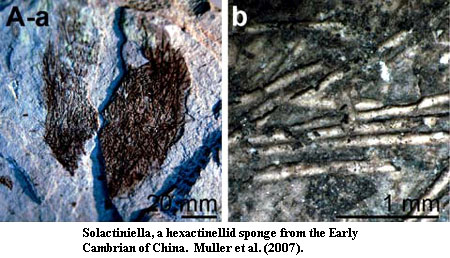
Fossil Record. However, the sponges clearly evolved in a very different direction from Fungi. One conspicuous specialization was the ability to make a mineralized skeleton. In fact, sponge spicules are so conspicuous that their absence from the Ediacaran fossil record seemed peculiar. Fortunately, clear examples have of hexactinellid, demosponge and, perhaps, calcarean sponges have now been recovered from the Ediacaran of Mongolia, China and India. Li et al. 1998); Tiwari et al. (2000).
Sponges and Eumetazoa. Not so long ago, the gap between sponges and Eumetazoa (corals + cows) seemed almost unbridgeable. But the distance between Porifera and Cnidaria has shrunk considerably. One recent molecular survey demonstrated that advanced sponges (Oscarella again) have not only a true epithelium, but essentially 100% of the eumetazoan complement of cell adhesion proteins, including cell-surface receptors, proteins for cytoplasmic bridges, and the proteins of the extracellular matrix. Nichols et al. 2006). What the sponges do not appear to have is a gut or a nervous system. These still appear to be synapomorphies of the Eumetazoa.
"Heteractinida": the Stem Group of all Sponges. In a recent, influential paper, Botting & Butterfield (2005) posit that all of the major sponge groups derived from the early "heteractinid" sponges, which they describe as follows: "Heteractinids are a problematic group of Paleozoic sponges characterized by hexaradiate spicules, a pattern not seen in extant forms, but with obvious symmetry relationships with the triradial spicules of calcareans; combined with the preserved carbonate mineralogy and bilaminar bilaminar structure of late Paleozoic forms, they have been widely accepted as calcareans."
 Based on a close examination of the Middle Cambrian Eifelia, they argue that at least some heteractinids had spicules containing both carbonate and siliceous layers. Certainly, Eifellia spicules had two layers, in addition to a core. What those layers were made of is less clear, but the speculation is at least reasonable. Without doubt, Eifellia had a peculiar mix of spicule symmetries. Its fossils include both flat, hexaradiate spicules (mostly large) and orthogonally symmetrical 4- or 6-armed spicules (mostly small). Thus, Botting & Butterfield assert that the heteractinellids were probably a heterogeneous group with a mosaic of characters we now think of as unique to one or another group of living sponges.
Based on a close examination of the Middle Cambrian Eifelia, they argue that at least some heteractinids had spicules containing both carbonate and siliceous layers. Certainly, Eifellia spicules had two layers, in addition to a core. What those layers were made of is less clear, but the speculation is at least reasonable. Without doubt, Eifellia had a peculiar mix of spicule symmetries. Its fossils include both flat, hexaradiate spicules (mostly large) and orthogonally symmetrical 4- or 6-armed spicules (mostly small). Thus, Botting & Butterfield assert that the heteractinellids were probably a heterogeneous group with a mosaic of characters we now think of as unique to one or another group of living sponges.
Botting & Butterfield (2005) deserves a close look, and is on our short list of Amazing Sponge Papers. It may be found at this direct link.
CONTINUED ON NEXT PAGE
page ATW070807
checked ATW070725,
last modified ATW080104
