
Ediacaran of Namibia. Grotzinger et al. (2000).


| Archaeocyatha | ||
| Porifera | Irregulares - 2 |
| Metazoa | Metazoa | Metazoa | ||||
| Time |
|
Porifera
├─Archaeocyatha
│ ├─Ajacicyathida
│ └─Irregulares
└─┬─Stromatoporoidea
└─┬─Calcarea
└─┬─Hexactinellida
└─┬─Demospongiae
│ ├─Tetractinomorpha
│ └─Ceractinomorpha
└─┬─Homoscleromorpha
└─Eumetazoa
|
Introduction |
| |
|
|
| D. Detailed texture of the internal "mesoclot" texture of many thrombolites. Note (a) the two different scales of structure and (2) the fact that the clots are composed of curved plates with a fairly consistent vertical axis. | E. Exterior of thrombolite. Note the pores on the exterior surface (brown) and, again the texture of small cells where the surface is abraded away (red). | |
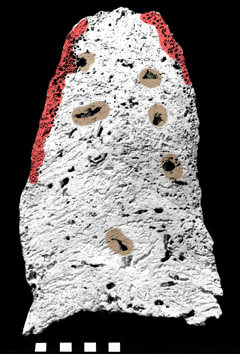
| Ediacaran of Namibia. Grotzinger et al. (2000). |
Recent of the Bahamas. Cyanobacterial Image Gallery. | |
| |
|
|
| F. Very common widening-upwards structure. | G. Thrombolite columns in cross-section. Note the peculiar structure in the upper left hand corner (blue) and, possibly, a similar structure on the lower right.. | |
| Unidentified thrombolite from Dupraz et al. (2006). | Unidentified thrombolites -- Recent of the Bahamas. From the Cyanobacterial Image Gallery | |
So, there you have it. Thrombolites, mostly simple cyanobacteria, can make all kinds of archaeocyath structures with no material help from hox genes or other body plan genetics. Thrombolites form hollow tubes, with interesting-looking mouths, external walls with pores, widening-upward cones, reticular systems of mineralized compartments, and mesoclot internal walls almost indistinguishable from the "thrombolitic" patterns of irregular archaeocyath septa. Sometimes, the walls appear to be doubled (not shown -- reference missing and presumed lost). We have not seen the cup-within-a-cup conformation as such, nor any convincingly tabulate structures, but neither of these features is present in the most basal archaeocyaths (Monocyathida), either. Thus every structure common to all archaeocyaths is also found in thrombolites, although there is no genetic commonality which can possibly explain the similarity.
So how do we pull the rabbit out of the hat?
The Automata of Dupraz sounds like a Twelfth Century treatise on alchemy, perhaps the work of a degenerate Florentine abbot, notorious for his unspeakable perversions. In fact, Dupraz et al. 2006) is an exceedingly Twenty-First Century paper, but somewhat magical for all that. The automata in the paper are not golems, but mathematical constructs which date back only to the work of John von Neumann. Dupraz and co-workers set out -- not to explain how cyanobacteria build such oddly complex structures -- but to see if they could model this behavior using a minimal set of mathematical rules.
Note that this is not paleobioloigy or paleoecology. Dupraz etc. do not try to guess what Neoproterozoic bacteria were actually like, or what their environment was. Theirs is simply a mathematical environment in which cellular automata move and form patterns according to a small set of rules. The rules imply no particular genetic program, no particular geochemistry, and no special ecological or trophic structure to the community -- just half a dozen rules. The rules contain parameters which can be varied. Those variations (on different runs, or in some pattern during runs) can mimic just about all general morphologies actually found in bacterial reefs. The parameters correspond, but only in a general way, to environmental constraints
If you're curious about details, see the link to this paper in the references section. The relevant point here is not whether this particular model is "correct" or sufficient. The point is that Neoproterozoic reef structures -- including the features shown in the images above -- probably do not require unusual circumstances or particular organisms. We don't try to explain the topology of a particular sandbox surface by examining the genomes of the children who have been playing in it. Other things are far more relevant, e.g. their numbers, ages, and the availability of buckets, water, and parental supervision.
Likewise it probably made little difference whether the biota of these microbialites was entirely cyanobacterial, or whether some eukaryotic protists or even very simple animals were thrown in. The morphology of the resulting microbialite would be much the same because it was largely controlled by factors unrelated to details of community structure. As long as the bulk of the material was calcimicrobial, nothing else would matter a great deal. This also survives an intuitive reality check. By the Late Neoproterozoic, microbialites had already been around for one or two billion years, possibly even longer. Eukaryotes had existed for a minimum of half a billion years and potentially far longer. Bacterial reefs had already survived and adapted to an enormous number of environmental and biotic challenges. We would not expect them to change much, and they have not, even today. Accordingly, it seems much more likely that the earliest metazoans on the reef would adapt to the existing furniture, rather than immediately redecorating.
So, our hypothesis is that early metazoans did not create body plans, so much as adapt to the pre-existing shapes of microbialites. Other than being physically possible, and parsimonious (since it does not require metazoans to invent body plans de novo), is there any evidence for this wild guess? Sort of. That is to say, it seems to be consistent with the very little we know about actual primitive metazoans. True, what we know is indeed little. Almost all of the Ediacaran animals are thought to be well off the main line of Metazoa, or outside Metazoa altogether. Worm-like trace fossils are not particularly helpful, either. They may represent the tracks of advanced, bilaterian animals. If so, they are irrelevant to the ancestry of sponges. Alternatively, some may represent the fossilized tubes of unknown things living below the surface. Whatever their relationships, we can know almost nothing about their morphology from the tubes alone.
This seems to leave two candidates, Cloudina and Namacalathus [7]. These two are often coupled in the literature, since they tend to be found in similar environments, i.e., on carbonate platforms and in association with cyanobacterial "build-ups." Almost all other animals from the Ediacaran are restricted to siliclastic sediments. Grotzinger et al. (2000). Cloudina and Namacalathus also have some degree of morphological similarity, although the resemblance is not particularly compelling. More interesting is the fact that these two animals, the only two regularly found on carbonate platforms are, probably by no coincidence, the only two Precambrian animals with carbonate mineralized skeletons.
 Cloudina was only lightly mineralized, apparently by the precipitation of calcite in an organic template (Grant 1990). Grotzinger et al. (2000) [6]. That may be helpful. In fact the usual reconstruction of Cloudina looks a little like the cone-within-a-cone structure of an archaeocyath. The difference is that the stack of cones keeps on growing, like a column of paper cups. Recently, this interpretation has been challenged. Miller (2004). However, Miller's reconstruction of Cloudina differs mainly in omitting the "test-tube bottoms." The resulting structure is a continuous calcite tube which periodically gives rise to rings of flange-like protrusions, but otherwise looks a bit like Figure F above. Miller's three-dimensional interpretation of these rings is that they are the externally visible sign of partially doubled or multiple walls, perhaps from repairs -- a bit like the arrangement in Figure C. If so, this is much more interesting. However, no one currently suggests that Cloudina had any of the intervallum intricacies that distinguish the archaeocyaths; and the actual living cells of an archaeocyath resided in the space between the two walls. Nevertheless, the presence of a partially double-walled, cup-shaped animal in precisely the environment which the archaeocyath would come to dominate a few million years later is, at least, a striking coincidence.
Cloudina was only lightly mineralized, apparently by the precipitation of calcite in an organic template (Grant 1990). Grotzinger et al. (2000) [6]. That may be helpful. In fact the usual reconstruction of Cloudina looks a little like the cone-within-a-cone structure of an archaeocyath. The difference is that the stack of cones keeps on growing, like a column of paper cups. Recently, this interpretation has been challenged. Miller (2004). However, Miller's reconstruction of Cloudina differs mainly in omitting the "test-tube bottoms." The resulting structure is a continuous calcite tube which periodically gives rise to rings of flange-like protrusions, but otherwise looks a bit like Figure F above. Miller's three-dimensional interpretation of these rings is that they are the externally visible sign of partially doubled or multiple walls, perhaps from repairs -- a bit like the arrangement in Figure C. If so, this is much more interesting. However, no one currently suggests that Cloudina had any of the intervallum intricacies that distinguish the archaeocyaths; and the actual living cells of an archaeocyath resided in the space between the two walls. Nevertheless, the presence of a partially double-walled, cup-shaped animal in precisely the environment which the archaeocyath would come to dominate a few million years later is, at least, a striking coincidence.
Another odd morphological similarity links Cloudina to sponges. The figure shows (A & B) part of the wall of Cloudina from Feng et al. (2002). C and D are cross-sections of a demosponge (Tethya) spicule taken from Weaver et al. (2003). The similarity is striking, despite an approximately 10-fold difference in scale. No doubt the structural resemblance could be due to the fact that there are only so many ways to build cylindrical objects from amorphous minerals. However, the possibility of homoplasy doesn't set us back.
Remember, we don't need to conclude that Cloudina and archaeocyaths are related. This is possible (see, e.g. Budd, 2003), but not necessary. Our point is that Cloudina and archaeocyaths both closely resemble thrombolites of various types. We are suggesting a common mode of evolution for sponges and Cloudina, not common ancestry. In fact, we have to be quite careful about trying to infer phylogenetic relatedness from morphology, if the more parsimonious interpretation is that morphology came before phylogeny. We'll worry about the mechanism for this peculiar reversal of the usual order in just a moment. For now, consider it simply as a reason not to jump to premature conclusions.
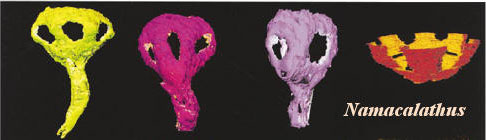 More or less contemporaneous with Cloudina is the tiny Namacalathus. If archaeocyaths are "ancient cups," then Namacalathus is an ancient goblet. The upper portion of Namacalathus is bowl-like, but with ovoid "port holes," usually six in number. In fact, Grotzinger et al. argue that Namacalathus actually had hexagonal symmetry. The bottom portion of the animal is relatively long and stem-like. At this point, it may be amusing to compare Grotzinger's reconstruction of Namacalathus with the ghostly outlines in the upper left-hand corner of Figure G above. This proves absolutely nothing, but it is interesting to contemplate that something similar to Namacalathus may have survived the end of the Proterozoic -- by over 540 My.
More or less contemporaneous with Cloudina is the tiny Namacalathus. If archaeocyaths are "ancient cups," then Namacalathus is an ancient goblet. The upper portion of Namacalathus is bowl-like, but with ovoid "port holes," usually six in number. In fact, Grotzinger et al. argue that Namacalathus actually had hexagonal symmetry. The bottom portion of the animal is relatively long and stem-like. At this point, it may be amusing to compare Grotzinger's reconstruction of Namacalathus with the ghostly outlines in the upper left-hand corner of Figure G above. This proves absolutely nothing, but it is interesting to contemplate that something similar to Namacalathus may have survived the end of the Proterozoic -- by over 540 My.
The principal non-thrombolite characteristic of Namacalathus is that, according to Grotzinger et al., the animal had a regular symmetry, specifically a more or less hexagonal symmetry about a central axis. This is particularly striking in cross-section. Extant sponges don't show this type of symmetry, nor do thrombolites. On the other hand, some basal archaeocyaths do show this pattern (Archaeolynthus [8], and perhaps Dokidocyathus, see Perejón & Moreno-Eiris, 2006) as do various blastula stage sponge embryos (Leys & Ereskovsky, 2006). In these latter cases, the symmetry is essentially an artifact, normally temporary and imposed by structural constraints during development. It is not the product of some hox-like genetic program. That is, early metazoan morphology was a matter of physical constraints, only later followed by genetic programming.
 So, based on these whirlwind generalizations, we seem to have a peculiar puzzle. We asked earlier, "How do we get the rabbit out of the hat?" Actually, getting the rabbit out is no problem. He'll eventually leave on his own. The real issue is how he got there in there in the first place. Magicians have only minutes, but the metazoans had megayears. We suggest that these particular rabbits were in their hats because they evolved there.
So, based on these whirlwind generalizations, we seem to have a peculiar puzzle. We asked earlier, "How do we get the rabbit out of the hat?" Actually, getting the rabbit out is no problem. He'll eventually leave on his own. The real issue is how he got there in there in the first place. Magicians have only minutes, but the metazoans had megayears. We suggest that these particular rabbits were in their hats because they evolved there.
The earliest metazoans had no particular shape, other than perhaps the generally spherical condition choanoflagellates have on the smallest scales. For such an organism, becoming trapped in a thrombolite on a bacterial reef would be an excellent thing. Surrounded by tasty bacteria, protected from environmental hazards -- who could ask for more? In this ideal environment, the major evolutionary pressures would be largely restricted to three factors: getting adequate circulation to receive food and remove waste, not outgrowing the space limitations, and dealing with environmental wear and tear on the thrombolite. Almost nothing else falls into the rather limited range of things which (a) matter to a small ball of cells and which (b) it could do anything about.
Very few of us know how to build a house from scratch; but almost everyone learns to make a few basic repairs. That requires much less skill. So, we suspect, the earliest metazoans had no expertise in general construction, but would gradually evolve the ability to make repairs by influencing the deposition of calcium carbonate, creating small holes for circulation, keeping the central region clear for the same reason, reinforcing the peripheral region, and ultimately creating a complete skeletal structure. The structure would, naturally, begin by looking more or less like a thrombolite. That is surely the most parsimonious expectation. The net result would ultimately be a collection of organisms which share some odd morphological characteristics with thrombolites, which are equipped with superficially similar biochemical toolkits for using calcium carbonate, but which do not seem to be very closely related to each other in other ways.
This leaves a great deal unexplained, but at least convinces us that there are reasonable explanations for what would otherwise be a remarkable series of odd coincidences at the base of the Metazoa and the dawn of the Phanerozoic. ATW070920.
ATW070920

{kind=link}