Homoscleromorpha - 2
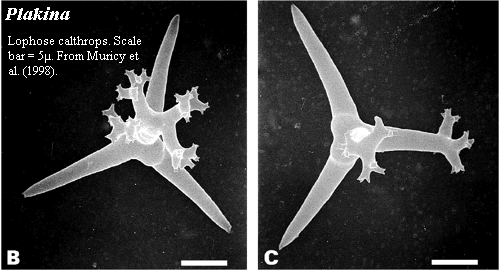 All homoscleromorphs have reduced skeletons, and some lack spicules entirely. When spicules are present, they are all generally of the same size -- hence the name homoscleromorph. Bergquist & Kelly (2004). Homoscleromorphs apparently do not have separate, sharply-defined size-classes of spicules. In this respect they differ from all three of the other main sponge groups.
All homoscleromorphs have reduced skeletons, and some lack spicules entirely. When spicules are present, they are all generally of the same size -- hence the name homoscleromorph. Bergquist & Kelly (2004). Homoscleromorphs apparently do not have separate, sharply-defined size-classes of spicules. In this respect they differ from all three of the other main sponge groups.
The homoscleromorph spicule complement includes silicate tetractines, just as in most demosponges. The homoscleromorph tetractine is of the type referred to as a calthrop. A "caltrop" or "calthrop" was an evil-looking medieval landmine, consisting of mainly of four short, sharpened spikes radiating from a common center at equal angles. These were scattered in front of defensive positions and covered under a thin layer of dirt, where they tended to inconvenience charging knights. Homoscleromorph tetractine spicules often have the same generally plan, albeit with considerably less military value. Fortunately, in practice, sponges are rarely called upon to receive the assaults of heavy cavalry, as such tactics tend to be ineffective in a marine setting.
As indicated in the image, the genus Plakina is characterized by having lophose calthrops -- calthrops with branched spines. Muricy et al. 1998).
 We have previously touched on the development of homoscleromorphs. There, we described it as generally similar to demosponge development. In retrospect, it seemed to us that we could have been just a tad more specific. So, although we can do little more than paraphrase Leys & Ereskovsky (2006), we supply some additional details here.
We have previously touched on the development of homoscleromorphs. There, we described it as generally similar to demosponge development. In retrospect, it seemed to us that we could have been just a tad more specific. So, although we can do little more than paraphrase Leys & Ereskovsky (2006), we supply some additional details here.
For a number of reasons, we do not normally talk much about reproductive biology
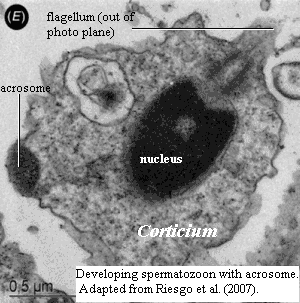
[19]. However, one frequently-mentioned character of homoscleromorphs falls into this category. The spermatozoa of many sexually reproducing eumetazoans bear an acrosome, the biochemical equivalent of an armor-piercing warhead to aid to fertilization. One of the folk-tales of spongeology is that only homoscleromorphs have acrosomes. Boute et al. (1996), Exposito et al. (2002), Leys & Ereskovsky (2006), Sperling et al. (2006). This belief appears to be mistaken. Acrosomes in calcareous sponges have been known for a long time. See, e.g., Gatenby (1927). Definitive evidence of acrosomes in demosponges has only recently been obtained. Riesgo, Taylor et al. 2007) (citing Riesgo & Maldonado, in press). Proacrosomal vesicles have been observed in yet other demosponges. Id.
As in the development of most sponges, cleavage of the fertilized egg starts out in an organized manner: holoblastic, equal, and synchronous. As in the development of almost everything else in the universe, matters quickly become disorganized, asynchronous and rather random. The result is an apparently haphazard solid ball of cells, the morula. Leys & Ereskovsky (2006). At this point, the details become a bit obscure -- at least to us -- but the essential moves are (a) rapid division of the cells on the surface, combined with (b) outward migration of cells in the interior. Ereskovsky & Gonobobleva (2000), Maldonado (2004), Leys & Ereskovsky (2006). These two maneuvers transform the solid morula into a hollow sphere of cells enclosing a cavity populated by symbiotic bacteria and collagen fibrils. Maldonado (2004). This method of forming a hollow ball of cells "multipolar egression") seems to be unique in the Metazoa. Leys & Ereskovsky (2006). It is often described as gastrulation. As previously discussed, we are not comfortable with that description.
 Within limits, the story so far may seems like a stripped down version of demosponge development. The main difference is that all migration is outward, rather than the complicated cell-sorting dance of the demosponges. Later events are even more like those occurring in demosponges:
Within limits, the story so far may seems like a stripped down version of demosponge development. The main difference is that all migration is outward, rather than the complicated cell-sorting dance of the demosponges. Later events are even more like those occurring in demosponges:
1) All, or almost all of the cells on the exterior become ciliated. As in demosponges, a small posterior region of the surface may be covered by larger cells which lack cilia. Maldonado (2004).
2) As in many demosponges the rapid division of the epithelial layer causes the surface to fold or wrinkle -- to become plicate. Ereskovsky & Gonobobleva (2000); Leys & Ereskovsky (2006).
3) The larva develops an equatorial belt of cells with a paracrystalline inclusions in their nucleus. Maldonado (2004). These cells may or may not be homologous to the light-sensitive belt of cells in demosponges.
4) The collagen in the internal cavity gathers under the epithelial layer to form a "felt." In homoscleromorphs, this structure becomes the basement membrane. Maldonado (2004).
5) The outline of the embryo becomes somewhat pear-shaped, as in demosponges and hexactinellids. Leys & Ereskovsky (2006).
After the larva settles, the posterior cells gradually overgrow the ciliated cells of the anterior surface and form the adult epithelium and pinacocytes. The blanketed anterior cells differentiate into the choanocytes and adult internal cells. Maldonado (2004). The belt of cells containing paracrystalline inclusions proliferate to form the excurrent canals the excurrent canal system. Id. In general, homoscleromorph metamorphosis resembles the same process in demosponges.
Relatively recent ultrastructural and biochemical work on homoscleromorphs shows that these sponges are surprisingly similar to non-sponge animals in characters relating to cell-cell communication and adhesion, such as the structure and extent of cell junctions and basic helix-loop-helix proteins, in addition to the famous basement membrane. Nichols et al. 2006); Simionato et al. (2007). As previously mentioned, comparative data are hard to come by. Consequently, these findings may suggest that Homoscleromorpha are close to the Eumetazoa, or simply that sponges are more similar to other animals than was once thought. Our bet is that both statements are correct.
The general feeling, at least as of this writing, seems to be that homoscleromorphs cannot be included in any of the three traditional groups of sponges. Until the late 1990's they were always included among the demosponges, and many workers still place them there. Even some RNA phylogenies tended to locate the clade within the demosponges, e.g., Shmitt (2003). However, most recent molecular phylogenies have nailed them outside the demosponges, either as demosponge sisters, or (perhaps more often) near-neighbors to the Calcarea (e.g. Sperling et al., 2006). The most sober and recent assessment is that neither morphology nor molecules have adequately resolved their relationship to the major sponge classes (Erpenbeck & Worheide, 2007); but no morphological study places the homoscleromorphs anywhere near the Calcarea. The two groups share no credible synapomorphies, while the homoscleromorphs have numerous similarities with demosponges.
On the other hand, both the molecular studies, comparative biochemistry, embryology, and ultrastructure all suggest that Homoscleromorphs have a special relationship to the Eumetazoa. Consequently, as argued here at really tiresome length, the most probable arrangement is that (a) homoscleromorphs diverged from near the base of the demosponges, and (b) this same branch ballooned into the Eumetazoa.
The homoscleromorphs have a miserable fossil record due to their reduced skeleton. They date back at least to the Mississippian. Unfortunately, they are not the only sponge group with calthrop spicules, and identification is difficult. They have probably never been numerous, and currently include only seven genera. Lehnert et al. 2005). Species are distinguished on the basis of texture, color, spicule size, branching pattern, and the structure of the water-canal system. Id.; Muricy et al. 1998). Not surprisingly, some of these identifications later turn out to be problematic. ATW080119
ATW080119
