

| Mollusca | ||
| Metazoa | Phylogeny |
| Metazoa | Mollusca | |||
| Annelida |
◄─┬─Aculifera
└─Conchifera |
Mollusca | Mollusca | Platyzoa |
|
Abbreviated Dendrogram
Spiralia
├─Brachiopoda
└─┬─Annelida
│
└─Mollusca
├─Aculifera
│ ╞═Mattheviidae
│ ├─Aplacophora
│ └─Polyplacophora
└─Conchifera
╞═Tryblidiida (Monoplacophora)
│ └─Cephalopoda
╘═Helcionelloida
╞═Rostroconchia
│ └─Scaphopoda
├─Bivalvia
│ ├─Protobranchia
│ └─Heterodonta
└─Gastropoda
╞═"Archaeogastropoda"
└─┬─Caenogastropoda
└─┬─Opisthobranchia
└─Pulmonata
|
The Ur-Mollusc Paradigm |
The following diagram presents a stylized relationship between the different Molluscan groups, with all evolving from a "hypothetical ancestral mollusk", which people consider to show shared ancestral characters for the group Mollusca
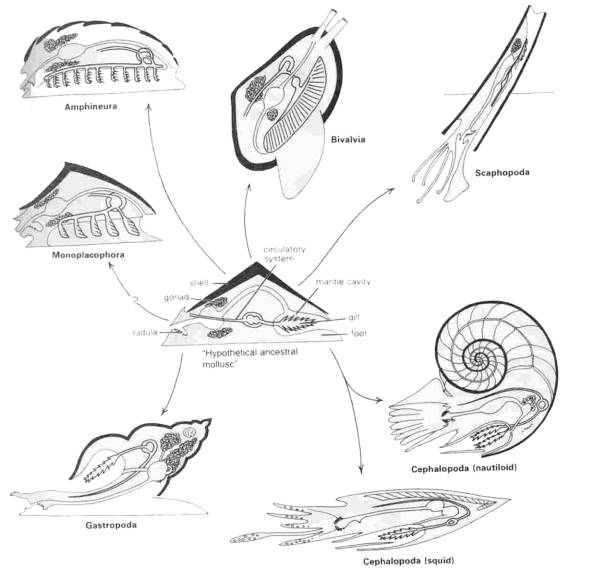
Useful as this diagram may be for illustrating the diversity among extant classes of mollusks, it really does nothing to explain the relationships of the various clades that make up the phylum. For this understanding, we must turn to the various hypotheses that have been proposed by paleontologists and neontologists over the last several decades. We can begin with an idea that has laid the foundations for much of our understanding of molluscan phylogeny, but has itself been later challenged - the Cyrtosoma-Diasoma hypothesis.
The view of molluscan evolution, which for some time assumed that the various classes developed from a hypothetical ancestral shelled ancestor, was revolutionized by an elegant model proposed by Bruce Runnegar and John Pojeta in 1974, and developed in a series of subsequent papers. This can be best represented by the following diagram:
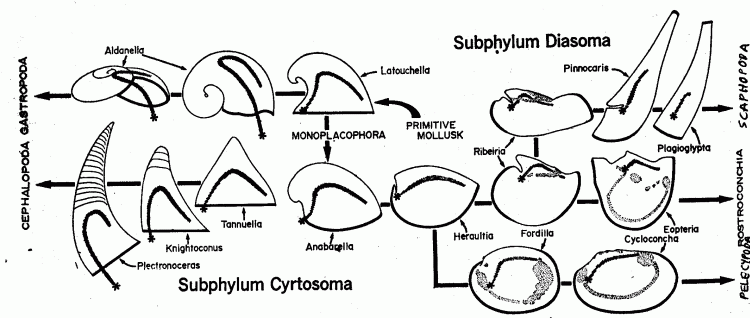
As indicated here, the Conchiferan (shell-bearing) mollusks are divided into two main sub-phyla, linked by a number of early transitional forms.
The Cyrtosoma (hunchback-body) includes the Monoplacophora, Gastropoda and Cephalopoda. The Monoplacophora gave rise to the Gastropoda, Cephalopoda, and the Rostroconchia (which latter is included in the second group). The Monoplacophora, Gastropoda, and Cephalopoda all are characterized primitively by a conical univalved shell, usually twisted into a spiral. The relatively small single aperture forces the anus to lie close to the mouth, and the gut is bent into a "U"-shape or (with gastropods) twisted on account of torsion. Most are surface-dwelling (epi-faunal) grazers or carnivores.
In the Diasoma (through-body) the gut is essentially straight. This group includes the classes Rostroconchia, Bivalvia and Scaphopoda, with the two latter derived from the Rostroconchia. These three classes form a lineage that diverged from the Monoplacophora in the Early Cambrian. They emphasized a shell form that in all groups is primitively open at both ends, allowing the gut to remain relatively straight, with an anterior mouth and posterior anus. They became burrowing (infaunal) deposit or filter feeders.
But although the Cyrtosomes - including as they do the Monoplacophora - are the ancestors of the Diasomes (and hence not strictly a monophyletic clade), both subphyla began to diversify within a few million years in the Early Cambrian.
Runnegar and Pojeta also suggest that the Polyplacophora can be derived from the Monoplacophora instead of (as usually suggested) a more primitive ancestral stock.
While not universally accepted, the Cyrtosoma-Diasoma paradigm presented an integrated paleontological perspective that complemented neontological discussions.
Yochelson (1978) rejects the Cyrtosoma-Diasoma model on the grounds that the differences between the shelled taxa remain too great, and that it would be foolish to speculate too much on the soft anatomy of animals known only from hard shells and a few rare muscle scars. Instead of a small number of classes linked by intermediate Cambrian shelled forms, he suggests a large number of different taxa that developed from a non-shelled ancestor or Archi-mollusk. This is represented in the following diagram
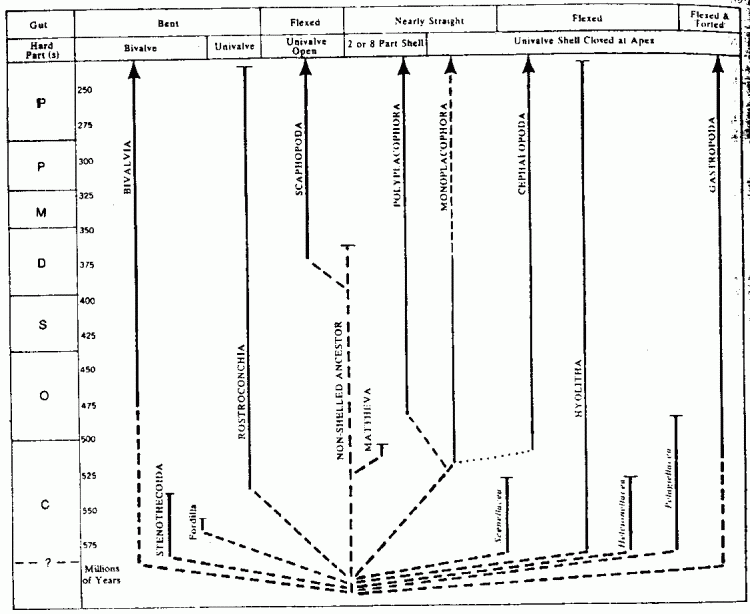
Here we are back to the original position at the top of the page, except that the ancestral form lacks a shell, and there are many early short-lived taxa that cannot be pigeonholed into the main classes.
This is reminiscent of the controversy between Gould and Conway-Morris' position regarding the Cambrian explosion and the number of extinct high level taxa. One sees a disorderly bush with a large number of separate early experiments, the other an orderly tree in which the branches can be carefully mapped out.
In 1972 C.R. Stasek theorized that the extant mollusks are the progeny of three separate lineages that separated before the phylum was well established. He stated no known fossil or living genera can bridge the gaps between any two of the three lineages, and treated each as a separate subphylum. These are (i) the Aculifera Hatscheck 1891, containing only the class Aplacophora, derived from the most primitive ancestor; (ii) the subphylum Placophora von Jhering 1876, containing only the class Polyplacophora, and emphasizing the pseudometamerism of its more advanced premolluscan ancestor; and (iii) the subphylum Conchifera Gegenbaur 1878, containing the remaining shelled mollusks - the Monoplacophora and the other classes derived from it.
Salvini-Plawen (1980) provided a reconsideration of the phylogenetic relationships among the various molluscan groups by studying various extant forms, including the little known soft-bodies vermiform types that had previously been called "Aplacophora" and lumped with the chitons under the now rejected class "Amphineura". On the basis of this analysis he proposed a phylogenetic line by which an original archi-mollusc gives rise to the main Conchiferan groups as well as a number of side-branches. All in all there are four stages of progressive differentiation. The first has an early side branch called the Scutopoda dividing from the main line, and eventually becoming the Caudofoveata (a class of Aplacophora). The other line is represented by the Adenopeda, which include the ancestors of all other mollusks. From the Adenapoda the shell-less Solenogastres (the second class of Aplacophora) separate from the shell-bearing Testaria. Within the Testaria the Polyplacophora, or "Placophora", divide from the Conchifera. Finally, the Conchiferan ancestor gives rise to the various extant classes, along with the extinct Galeroconcha (Monoplacophora plus Bellerophontida, the latter here considered to be untorted). The Aplacophora are rejected, as the Placophora and Solenogastres are tied together by a number of shared unique characteristics (synapomorphies), in contrast to the merely symplesiomorphic (shared primitive) characters in Solenogastres and Caudofoveata. This can be represented by the following diagram:
|
|
|
| "Diagram of the phylogenetic radiation of me Mollusks (black bars indicate fossil records, time-scale in millions of years, logarithmic). (1) Hypothetical archi-mollusc (main organization. and ventral view) with overall ventral gliding surface (black), postero-lateral mantle cavity, scale-bearing mantle, straight midgut pouched laterally, serial dorsoventral muscle bundles, gonopericardial system. and main nervous system with terminal sense organ(s). (2) Evolutionary branch of burrowing Scutopoda (lateral and ventral) with cerebrally-innervated section of locomotor surface (= pedal shield, black) and reduction of its ventrally- innervated section. (3) Evolutionary branch of gliding-creeping Adenopoda (lateral and ventral) with ventrally-innervated section of locomotor surface (= foot, black) including the pedal gland, with rudiment of head, and with peripedal-preoral mantle cavity. (4) Level of primitive Heterotecta (dorsal view), dorsal mantle with seven transverse rows of juxtaposed scaly bodies. (5) Level of Solenogastres (ventral and dorsal) with narrowed body and toot, mantle cavity reduced to preoral sensory pit (atrial sense organ) and to internal tubes (spawning ducts); adult mantle cover altered again to homogeneous arrangement of scales (6) Level of early Placophora (= Heptaplacota; dorsal view) with consolidation of juxtaposed scaly bodies to seven shell plates. (7) Regressive dorsoventral musculature in Caudofoveata. (8) Serial arrangement of dorso- ventral musculature in Solenogastres; compare (1). (9) Serial arrangement of dorso- ventral musculature in recent Placophora, concentrated according to the eight shell plates (8 x 2 = 16). (10) Primitive Conchifera with further concentration of the placophoran dorso-ventral musculature (9) according to the homogeneous conchs (see Neopilina)" - images and text © Institute of Malacology and Field Museum of Natural History, from Salvini-Plawen 1980 figure p. 258, caption page 259 | |
The above represents what might be called the Adenopoda hypothesis. This is complimentary to the Cyrtosoma-Diasoma hypothesi, the former pertaining to the most primitive molluscs, the latter to more advanced types.
The alternative to the Adenopoda hypothesis is the Aculifera hypothesis. This is simpler than Stasek's original hypothesis as it has only two subphyla, the Aculifera (spiny mollusks) and the Conchifera (shell bearing mollusks).
The subphylum Aculifera comprises the most primitive group - all those mollusks that do not produce a complete shell. Instead they have a hard exterior skin called a cuticula. This cuticula additionally is protected by calcareous spines (spiculi). Two classes belong to the Aculifera:
The Conchifera includes those mollusks that have a complete shell (although this may be lost in some more derived forms). Included here are the gastropods, bivalves, cephalopods, and other lesser groups, as follows:
The Visceroconcha containing the classes Gastropoda and Cephalopoda, and the Loboconcha containing the classes Scaphopoda and Bivalvia, are clearly the neontological equivalents to Runnegar and Pojeta's Cyrtosoma and Diasoma [ Mizzaro-Wimmer and Salvini-Plawen 2001]
John S. Peel reinterpreted the Cyrtosoma and Diasoma by rejecting the old "wastebasket" taxon Monoplacophora, and replacing it with two distinct and only distantly related early classes. These are the exogastrically-curved Class Tergomya (the superfamily Tryblidiacea), based upon a subclass of the same name described by Horny (1965a, b) and including the order Tryblidiida with the recent Neopilina; and a group of endogastrically coiled univalved mollusks, the Class Helcionelloida (the former superfamily Helcionellacea), including a range of primitive extinct firms, including some with cap shaped shells, others with high shells, even forms with "snorkels". The Helcionelloida is considered to be the direct ancestor of the Class Rostroconchia and members of this latter class are also interpreted here as endogastric mollusks. The model of diasome evolution proposed by Runnegar & Pojeta is replaced by one in which two parallel lineages are recognized within the former Diasoma: Tergomya - Bivalvia, and the Helcionelloida - Rostroconchia.
The scaphopods are tentatively derived from the Rostroconcha (although they may come from either lineage), and the Cephalopoda from the helcionelloids rather than from high-shelled conical Tergomya such as Knightoconus [Yochelson et al 1973). This, together with the derivation of the Gastropoda from the Tergomya or tergomyan-like forms, suggests that the Cyrtosoma is also not monophyletic.
During the 90s the two competing hypotheses of evolutionary relationships among conchiferan molluscs: the Diasoma-Cyrtosoma concept recognizing the clades (Tryblidiida, (Bivalvia, Scaphopoda), (Gastropoda, Cephalopoda)); and the Helcionellid concept with (Tryblidiida, (Bivalvia, (Gastropoda, Scaphopoda, Cephalopoda))). The rostroconches were either stem group diasomes or stem group scaphopods (Runnegar & Pojeta, 1974; Salvini-Plawen, 1980; Steiner, 1992; Engeser & Riedel, 1996; Runnegar, 1996); if the latter the Bivalvia stood alone as a monophyletic clade.
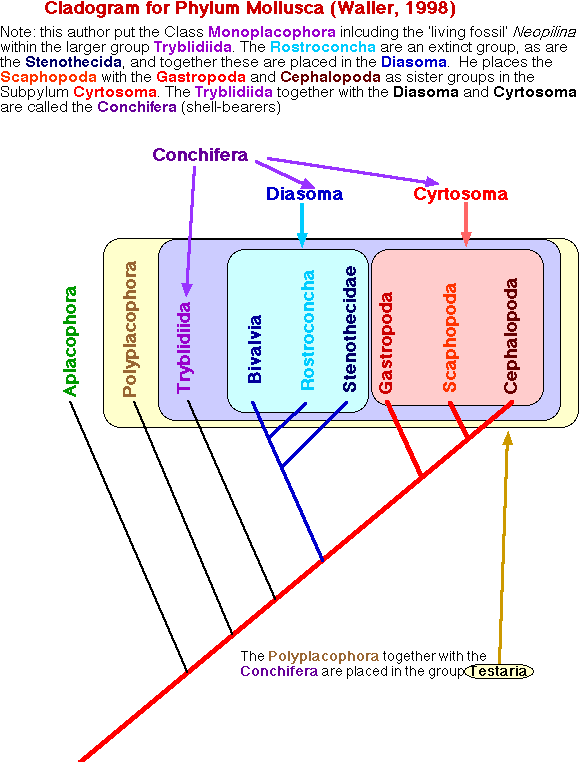 image and text by Ellen Thomas - from Biological Diversity and Evolution through Time [http: ethomas. web. wesleyan. edu/ scie639/ bivalve.htm] image and text by Ellen Thomas - from Biological Diversity and Evolution through Time [http: ethomas. web. wesleyan. edu/ scie639/ bivalve.htm] |
Both of these hypotheses have problems. The Diasoma results in convergent evolution of elongated dorso-ventral body axes in Scaphopoda and Cyrtosoma (Gastropoda, Cephalopoda), of multiple cephalic tentacles and ring-shaped muscle attachment in Scaphopoda and Cephalopoda. The Helcionellid concept has to account for convergent similarities in the development of the mantle-shell, the typical burrowing foot, and the nervous system in Bivalvia and Scaphopoda. [Steiner and Dreyer 2002]
Waller (1998) proposed a third alternative, that the Scaphopoda and Cephalopoda are sister groups within the subphylum Cyrtosoma. Waller's cladogram is shown below.
This established the new view by which extant conchiferan classes are grouped as follows (Tryblidiida, (Bivalvia, (Gastropoda, (Scaphopoda, Cephalopoda)))). The following represents a diagramatic interpretation of the Cyrtosoma - Diasoma hypothesis.
Steiner and Dreyer [2002] used molecular analysis and a 18S rDNA data set containing 17 Bivalvia, 6 Gastropoda, 17 Scaphopoda, 4 Cephalopoda, and 3 Polyplacophora. All the resulting trees showed Cephalopoda and Scaphopoda as sister groups with the Gastropoda connecting to their common stem. So far this supports Waller. However Bivalvia here appear paraphyletic at the base of the conchiferan ingroup. This is perhaps more a problem with the unreliability of current molecular phylogeny, due to "long branch attraction", than an actual representation of the early history of the group.
In 2000, Gerhard Haszprunar argued that the two aplacophoran taxa, the Solenogastres and Caudofoveata do not form a single clade. Based on current knowledge and available data, he concluded that the arrangement (Solenogastres (Caudofoveata (Polyplacophora + Conchifera))) is the most parsimonious one, with the Aplacophora as a basic, paraphyletic assemblage. The monophyly of the Testaria (Polyplacophora + Conchifera) is well supported by several characters, and the Aculifera (Polyplacophora + Aplacophora) rejected. The Adenopoda (Solenogastres + Testaria) of Salvini-Plawen is shown to be polyphyletic in this interpretation.
Further insights have been provided by the discovery of the Silurian spiny worm-like soft-bodied plated mollusk Acaenoplax hayae, which combines characteristics of both the Caudofoveata and the Polyplacophora, and can be seen as a sister clade to the Caudofoveata, and a sort of missing link or transitional form between these two groups [Sutton et al 2001], although this interpretation has been challenged by Steiner and Salvini-Plawen 2001, who claim Acaenoplax is a polychaete worm; an interpretation that is unlikely if only because of the similar number of transverse plates and bristles in both Acaenoplax, chitons, and larval Solenogastres, implying a close relationship between all these groups.
Steiner and Dreyer 2002 suggest a scenario for the evolution of the higher conchiferan taxa. Acquiring an infaunal life style occurred independently in the early Rostrochonchia-Bivalvia lineage and, later, in Scaphopoda, rendering the relevant characters convergent. After the separation of the gastropod lineage, the stem-group of Scaphopoda and Cephalopoda comprised high-shelled, dorsoventrally elongated helcionellids with a deep mantle cavity, multiple cephalic feeding tentacles, and a circular area of muscle-shell connection. Shell-chambering for buoyancy then led to the first cephalopods, and anterior mantle elongation and transient forms with posterior shell slits or snorkels in response to an increasingly infaunal habit gave rise to the tube-shelled scaphopods.
Thus, all rostroconches and some helcionellacean univalves technically become stem group bivalves. This is because the total group is thought to have originated at the latest common ancestor of the Bivalvia and the Cyrtosoma; Cytosoma here being (Gastropoda (Cephalopoda, Scaphopoda)). [Runnegar 2002].
While molluscan phylogeny is mostly concerned with extant, well-known taxa, as these can provide the most data, this avoids the fact that during the early Paleozoic molluscs were far more diverse than they are now, and included a large number of types that don't fit into the crown group taxa, and which for this reason provide useful phylogenetic data. The comparison of the various hypotheses as regards the status of various little known fossil groups. as well as better known extant classes, can be shown as follows
| Hypothesis | Runnerger & Pojeta | Yochelson | Peel | Waller | Wagner | Steiner and Dreyer |
| Conchifera evolution | Cyrtosoma-Diasoma | Archi-mollusk | Helcionellid-Tergyma | Cyrtosoma-Diasoma | -- | -- |
| Hyolitha | extinct related phylum | extinct class of Molluscs | -- | -- | -- | -- |
| extinct classes | Rostroconchia only | at least three and probably more | Rostroconchia and Helcionelloida | Rostroconchia only | -- | -- |
| Mathevia | polyplacophoran | extinct class | -- | -- | -- | -- |
| Monoplacophora | Highly diverse | Restricted to a few forms | Restricted to a few forms (Tergmya) | -- | -- | -- |
| Stenothecoides | bivalved monoplacophoran | extinct class | extinct class | -- | -- | -- |
| Helcionellida | Monoplacophora | probably extinct class | extinct class | stem grade | stem grade | stem grade |
| Rostroconcha | Diasoma | archi-mollusk derived | Helcionellid-derived | Diasoma - Stenothecidae derived | Stenothecidae derived | Helcionellid-derived |
| Bivalvia | Diasoma - Rostroconcha derived | archi-mollusk derived | Tergmya derived | Diasoma - Rostroconcha sister group | Rostroconcha sister group | Rostroconcha derived |
| Fordilla | Diasoma | extinct class | -- | Early Bivalve | Early Bivalve | -- |
| Gastropoda | Cyrtosoma | archi-mollusk derived | Tergmya derived | -- | "Tergmya" derived | Cyrtosoma |
| First Gastropods | Early Cambrian | Furongian | Furongian | -- | Furongian | -- |
| Aldanella | early gastropod | not a mollusk | -- | -- | -- | -- |
| Pellagiellacea | Early Gastropod | possibly extinct class | -- | -- | Paragastropod | -- |
| Scaphopoda | Diasoma - Rostroconch derived | archi-mollusk derived | Helcionellid Rostroconch derived | Cyrtosoma - sister group Cephalopods | -- | Helcionellid derived - sister group to Cephalopods |
| first Scaphopoda | Mid-Ordovician | Devonian or Early Carboniferous | -- | -- | -- | -- |
| Cephalopoda | Cyrtosoma - Hypseloconid derived | archi-mollusk derived | Helcionellid derived | Cyrtosoma - sister group Scaphopoda | -- | Helcionellid derived - sister group Scaphopoda |
By the start of the 21st century, morphology-based cladistics and molecular phylogeny had resulted in a number of conflicting hypotheses, some more popular than others. Some of these are summarised in the following diagram.
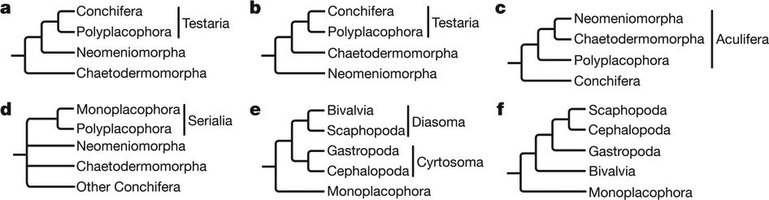 Leading hypotheses of molluscan phylogeny, from Kocot et al 2011, references from Kocot & Halanych 2009. a, Adenopoda hypothesis placing Chaetodermomorpha basal (Salvini-Plawen 1972, 1980, 1985, 1991, 2003). b, Hepagastralia hypothesis placing Neomeniomorpha basal (Salvini-Plawen and Steiner 1996). c, Aculifera hypothesis placing Aplacophora sister to Polyplacophora (Ivanov 1996, Scheltema 1993, 1996, Kocot et al 2011, Smith et al 2011), modified version with Polyplacophora ancestral to Aplacophora (Sigwart & Sutton 2007, Vinther et al 2011). d, Serialia hypothesis allying Polyplacophora and Monoplacophora (Giribet et al 2006). e, Diasoma and Cyrtosoma hypotheses allying bivalves to scaphopods and gastropods to cephalopods, respectively (Runnegar and Pojeta 1974, Salvini-Plawen 1980, Simone 2009). f, Unnamed hypothesis, allying scaphopods and cephalopods (Giribet and Wheeler 2002, Haszprunar 2000, Passamaneck et al. 2004, Steiner and Dreyer 2002, 2003, Waller 1998). (not figured) Conchifera, grouping together Monoplacophora, Gastropoda, Bivalvia, Cephalopoda, and Scaphopoda (Wingstrand 1985) and Testaria, with Polyplacophora (but not aplacophora) sister to Conchifera (Salvini-Plawen 1972, 1980, 1985, Salvini-Plawen and Steiner 1996, Haszprunar 2000.), unnamed clade grouping Scaphopoda, Gastropoda, Bivalvia (Smith et al 2011), unnamed clade with Monoplacophora and Cephalopoda as sister taxa (Smith et al 2011), and Pleistomollusca allying Gastropoda and Bivalvia (Kocot et al 2011) Leading hypotheses of molluscan phylogeny, from Kocot et al 2011, references from Kocot & Halanych 2009. a, Adenopoda hypothesis placing Chaetodermomorpha basal (Salvini-Plawen 1972, 1980, 1985, 1991, 2003). b, Hepagastralia hypothesis placing Neomeniomorpha basal (Salvini-Plawen and Steiner 1996). c, Aculifera hypothesis placing Aplacophora sister to Polyplacophora (Ivanov 1996, Scheltema 1993, 1996, Kocot et al 2011, Smith et al 2011), modified version with Polyplacophora ancestral to Aplacophora (Sigwart & Sutton 2007, Vinther et al 2011). d, Serialia hypothesis allying Polyplacophora and Monoplacophora (Giribet et al 2006). e, Diasoma and Cyrtosoma hypotheses allying bivalves to scaphopods and gastropods to cephalopods, respectively (Runnegar and Pojeta 1974, Salvini-Plawen 1980, Simone 2009). f, Unnamed hypothesis, allying scaphopods and cephalopods (Giribet and Wheeler 2002, Haszprunar 2000, Passamaneck et al. 2004, Steiner and Dreyer 2002, 2003, Waller 1998). (not figured) Conchifera, grouping together Monoplacophora, Gastropoda, Bivalvia, Cephalopoda, and Scaphopoda (Wingstrand 1985) and Testaria, with Polyplacophora (but not aplacophora) sister to Conchifera (Salvini-Plawen 1972, 1980, 1985, Salvini-Plawen and Steiner 1996, Haszprunar 2000.), unnamed clade grouping Scaphopoda, Gastropoda, Bivalvia (Smith et al 2011), unnamed clade with Monoplacophora and Cephalopoda as sister taxa (Smith et al 2011), and Pleistomollusca allying Gastropoda and Bivalvia (Kocot et al 2011) |
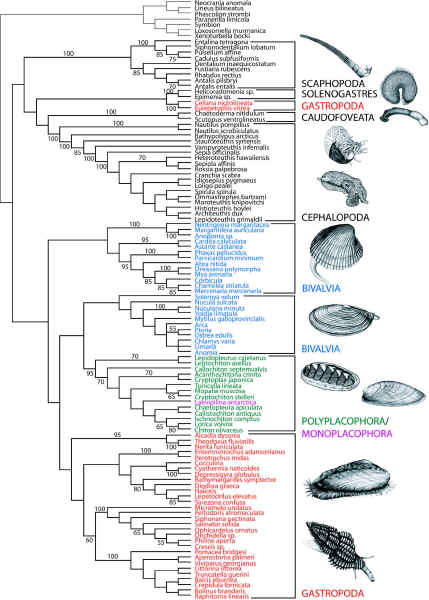 Giribet et al 2006 fig.2. Monoplacophora is nested within the Polyplacophora. Bivalvia emerges as a paraphyletic group. Scaphopoda, Solenogastres (Neomeniomorpha ), Caudofoveata (Chaetodermomorpha), and the Cephalopoda make a monophyletic clade. (via Taxa of Life) |
Over the past decade or so, developments in both molecular phylogeny and cladistic methodology have if anything only obscured rather than contributed further insights, with lack of clear consensus between morphology and molcules, and even different molecular phylogenies The following two paragraphs are taken from Wikipedia, tending a more comprehensive coverage on our part.
"It is uncertain whether the ancestral mollusc was metameric (composed of repeating units) (which would suggest an origin from an annelid-like worm (Jacobs et al 2000). According to Giribet et al 2006 that the repetition of gills and of the foot's retractor muscles were later developments, while Sigwart & Sutton 2007 concluded that the ancestral mollusc was metameric, and that it had a foot used for creeping and a "shell" that was mineralized. In one particular one branch of the family tree, the shell of conchiferans is thought to have evolved from the spicules (small spines) of aplacophorans; however this is difficult to reconcile with the embryological origins of spicules (Henryetal2004).
The molluscan shell appears to have originated from a mucus coating, which eventually stiffened into a cuticle. This would have been impermeable and thus forced the development of more sophisticated respiratory apparatus in the form of gills. Eventually, the cuticle would have become mineralized (Runnegar & Pojeta 1974, using the same genetic machinery (engrailed) as most other bilaterian skeletons (Jacobs et al 2000). The first mollusc shell almost certainly was reinforced with the mineral aragonite (Porter 2007).
Morphological analyses tend to recover a conchiferan clade that receives less support from molecular analyses (Winnepenninckx et al 1996), although these results also lead to unexpected paraphylies, for instance scattering the bivalves throughout all other mollusc groups (Passamaneck et al 2004).
However, an analysis in 2009 that used both morphological and molecular phylogenetics comparisons concluded that the molluscs are not monophyletic; in particular, that Scaphopoda and Bivalvia are both separate, monophyletic lineages unrelated to the remaining molluscan classes—in other words that the traditional phylum Mollusca is polyphyletic, and that it can only be made monophyletic if scaphopods and bivalves are excluded (Goloboff et al 2009). A 2010 analysis managed to recover the traditional conchiferan and aculiferan groups, but similarly concluded that the molluscs are not monophyletic, this time suggesting that solenogastres are more closely related to the non-molluscan taxa used as an outgroup than to other molluscs (Wilson et al 2010). Current molecular data is insufficient to constrain the molluscan phylogeny, and since the methods used to determine the confidence in clades are prone to over-estimation, it is risky to place too much emphasis even on the areas that different studies agree (Wägele et al 2009. Rather than eliminating unlikely relationships, the latest studies add new permutations of internal molluscan relationships, even bringing the conchiferan hypothesis into question (Vinther et al 2011)." -- Wikipedia
Some of these problems can be explained as due to this being still early days of molecular phylogeny, others to the fact that original molluscan evolution was part of the Cambrian explosion which involved a very rapid radiation of new lineages, and that molecular sequencing does not have the resolution to unravel the particular order of branching and new clades (this is the same problem as with metazoa in general). It is possible however that better sampling and more broad ranging studies will improve resolution. In 2011, two deep phylogeny analyses produced a molecular consensus emerging totally at odds with both morphology and paleontology; for example bivalves and gastropods as sister taxa. Pending a more comprehensive review, the cladograms each produce are shown here:
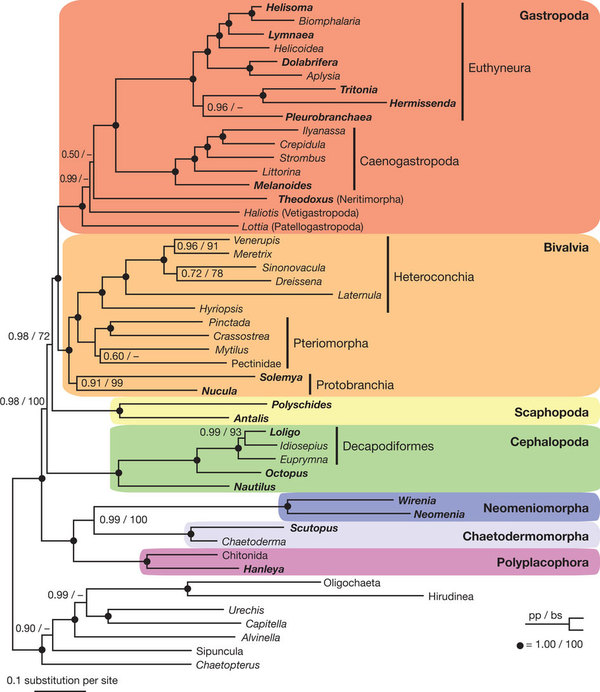
From Kocot et al 2011 (abstract below) and phylogram (right):
"Evolutionary relationships among the eight major lineages of Mollusca have remained unresolved despite their diversity and importance. Previous investigations of molluscan phylogeny, based primarily on nuclear ribosomal gene sequences or morphological data, have been unsuccessful at elucidating these relationships. Recently, phylogenomic studies using dozens to hundreds of genes have greatly improved our understanding of deep animal relationships. However, limited genomic resources spanning molluscan diversity has prevented use of a phylogenomic approach. Here we use transcriptome and genome data from all major lineages (except Monoplacophora) and recover a well-supported topology for Mollusca. Our results strongly support the Aculifera hypothesis placing Polyplacophora (chitons) in a clade with a monophyletic Aplacophora (worm-like molluscs). Additionally, within Conchifera, a sister-taxon relationship between Gastropoda and Bivalvia is supported. This grouping has received little consideration and contains most (>95%) molluscan species. Thus we propose the node-based name Pleistomollusca. In light of these results, we examined the evolution of morphological characters and found support for advanced cephalization and shells as possibly having multiple origins within Mollusca."
The phylogram can be simplified to:
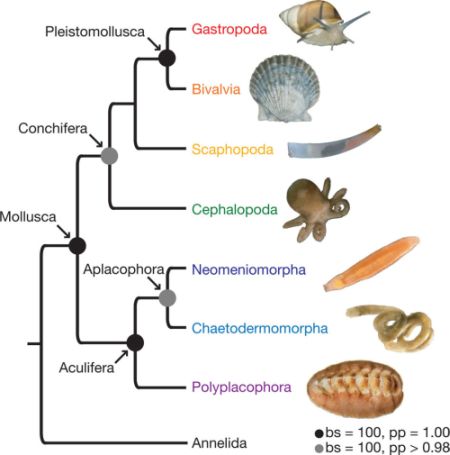 Deep molluscan phylogeny as inferred by Kocot et al 2011. Black circles represent nodes with bs = 100 and pp = 1.00. Gray circles represent nodes with bs = 100 and pp ≥ 0.98. |
Both this phylogeny and the following support both the Aculifera (aplacocophara + polyplacophora) and the Conchifera. Both place the cephalopods in a basal position, and a similar result was recovered by Giribet et al 2006. The implication being that the advanced nervous system found in both cephalopods and gastropods evolved by convergence, as did similiar morphological features between cephalopods and scaphopods. Smith et al 2011 (below) make cephalopods and monoplacophorans sister taxa, which would support Cambrian fossils like Knightoconus, Hypseloconus, and/or Tanuella as intermediate forms, and a "monoplacophoran" origin is proposed also on a total evidence approach using fossil and morphological as well as molecular findings by Kröger et al 2011. Rather more surprising is that both affirm a clade consisting of bivalves, gastropods, and scaphopods, as either scaphopoda + (gastropoda + bivalvia) (Kocot et al 2011, also found by Vinther et al 2011), or bivalvia + (gastropoda + scaphopoda) (Smith et al 2011). It's hard to get a more opposite result to both morphology (which even in recent cladistic analysis (Simone 2009), paleontology, and ecological functionalism, all of which both favour the Diasoma-Cyrtosoma hypothesis. With such strong inconguuancy between morphology and molecules the possibility of sequencing artifacts such as long chain attraction (as seems to be the case when turtles are resoved as (very unlikely) archosaurs) makes more sense to us than the number of reversals and convergences required to force morphology into a molecular tree. Of course this is the old problem of which methodology carries the more reliable phylogenetic signal. Here we need to look at fossils of transitional forms, morphology, ecology, and evo-devo development as well as molecules.. Regarding fossil groups, Rostroconchia might be a candidate for a transitional group between bivalves and scaphopods, although where gastropods fit is anybody's guess. MAK120504

From Smith et al 2011 (abstract below) and phylogram (right):
"Molluscs (snails, octopuses, clams and their relatives) have a great disparity of body plans and, among the animals, only arthropods surpass them in species number. This diversity has made Mollusca one of the best-studied groups of animals, yet their evolutionary relationships remain poorly resolved. Open questions have important implications for the origin of Mollusca and for morphological evolution within the group. These questions include whether the shell-less, vermiform aplacophoran molluscs diverged before the origin of the shelled molluscs (Conchifera) or lost their shells secondarily. Monoplacophorans were not included in molecular studies until recently, when it was proposed that they constitute a clade named Serialia together with Polyplacophora (chitons), reflecting the serial repetition of body organs in both groups. Attempts to understand the early evolution of molluscs become even more complex when considering the large diversity of Cambrian fossils. These can have multiple dorsal shell plates and sclerites or can be shell-less but with a typical molluscan radula and serially repeated gills. To better resolve the relationships among molluscs, we generated transcriptome data for 15 species that, in combination with existing data, represent for the first time all major molluscan groups. We analysed multiple data sets containing up to 216,402 sites and 1,185 gene regions using multiple models and methods. Our results support the clade Aculifera, containing the three molluscan groups with spicules but without true shells, and they support the monophyly of Conchifera. Monoplacophora is not the sister group to other Conchifera but to Cephalopoda. Strong support is found for a clade that comprises Scaphopoda (tusk shells), Gastropoda and Bivalvia, with most analyses placing Scaphopoda and Gastropoda as sister groups. This well-resolved tree will constitute a framework for further studies of mollusc evolution, development and anatomy."

A consensus phylogeny, incorporating fossil forms, might be something like the following diagram by Telford & Budd (2011 (left)
There is actually some paleontological support for this new phylogeny as regards the Conchifera (right side of diagram), for example some high-shelled monoplacophorams ap[pear to be transitional or on the road to nautiloids (early shelled cephalopods) (Yochelson et al 1973, Webers & Yochelson 1989, Kröger et al 2011), whilst scaphopods, bivalves, and gastropods can all be derived from infaunal and epifaunal Helcionellids.
More problematic is the monophyletic Aculifera (left side of diagram). Despite both molecular and and paleontological-statistical cladistic data (Sigwart & Sutton 2007, Vinther et al 2011), there are no aculiferan synapomorphies, other than plesiomorphic halwaxiid (stem molluscan or basal spiralian) characteristics such as spicules and multiple dorsal shell plates. A monophyletic aculifera also leaves the ancestry of the Conchifera hanging in the air, as there are no other molluscan forms intermediate between the halwaxiids and crown group conchiferans. Along with some uncertainty regarding the status of the most primitive extant aculifera such as the worm-like solenogastres, and neontological morphology which strongly supports a paraphyletic aculifera, there is the possibility of molecular phylogeny showing a "false monophyly", as with the morphologically exceedingly unlikely cyclostomata clade of lampreys+ hagfish. A paraphyletic aculifera hypothesis is weakened by the fact that no transitional forms bewteen aculiferans and conchifera are known, although Giribet et al (2006)'s much disputed (on morphological grounds) Serialia could bridge the gap, if it is a paraphyletic grade rather than a monophyletic clade. It is quite possible also that the Aculiferan-Conchiferan split occurred very early, and that some soft-bodied forms like the annelid- and aplacophora-like Acaenoplax may represent a late (Silurian) survivor stem lineage ancetsral to both. Alternatively the pine-cone like Turrilepadids may be intermediate forms between the spiny and armoured halwaxiids and the molluscs in general. Initial cladistic analysis (Sigwart & Sutton 2007) show the turrilepadids to be stem-aplacophora, although this could change with further analyses and new material. For now we have tentatively assumed a paraphyletic aculifera leading to a monophyletic conchifera MAK120607
page by M. Alan Kazlev
checked ATW050713
page uploaded 28 December 2002, revised MAK120605, Creative Commons Attribution