| Palaeos | Sauropterygia | |
| Vertebrates | Neusticosaurus |
| Page Back | Unit Home | Unit Dendrogram | Unit References | Taxon Index | Page Next |
| Unit Back | Vertebrates Home | Vertebrate Dendrograms | Vertebrate References | Glossary | Unit Next |
|
Abbreviated Dendrogram
DIAPSIDA |--ARCHOSAUROMORPHA | LEPIDOSAUROMORPHA |--SAUROPTERYGIA | |--Placodontia | |--Wumengosaurus | |--Pachypleurosauridae | | |--Hanosaurus | | `--+--+--Dactylosaurus | | | `--Anarosaurus | | `--Serpianosaurus--Neusticosaurus-- pusillus--N. peyeri--N. edwardsii | `--+--Nothosauridae | `--Plesiosauria | |--Pliosauroidea | `--Plesiosauroidea | |--Elasmosauridae | `--Cryptocleidoidea `--Lepidosauriformes |--SPHENODONTIA `--SQUAMATA |
Contents
Overview |
Palaeos com, like other websites that take as their topic the evolutionary history of life, presents the relationship of life past and present in terms of tree like diagrams, called dendrograms. Each dendrogram is based on cladistic analyses and other studies; it represents an amalgamation of one or more phylogenetic hypotheses, represented in diagramatic form as cladograms (although dendrograms and cladograms are confused, most of what you read on the web that purports to be a cladogram is actually a dendrogram). Now, the whole thing about cladograms is that they are hypotheses for testing different phylogenies (geneologies of life so to speak). They don't actually tell you who the ancestors are, or even anything about those ancestors. All they do is provide statistical tests, on the basis of the data keyed in, on which species are most likely to be related to each other (these are called sister groups). This might seem a strange attitude to take, but the simple fact is that the fossil record, the trace of past life, is incomplete, and that the further back in deep time one goes, the more incomplete it becomes. Sometimes however things are fortuituous enough that we can get a glimpse of evolution in action, and see who the ancestors are. One such instance involves observation of a stratigraphic succession of pachypleurosaurs from the Middle Triassic of the Monte San Giorgio region, Switzerland (O'Keefe & Sander 1999). Deposition was continuous, and pachypleurosaurs common. Over a period of perhaps five million years (not long by geological standards, equivalent to Plio-Pleistocene or Quaternary time) these creatures lived and died and left their fossilised remains in the sediment. By comparing large numbers of specimens, it was possible to observe a single sequence of gradational evolution (this is called anagenesis), involving the series Serpianosaurus mirigiolensis, Neusticosaurus pusillus, N. peyeri, and finally N. edwardsii. The phylogeny was reconstructed using a combination of cladistic, phenetic, and stratigraphic (fossils and fossil sequences) information and methodologies. The result reveals the way that nature does not follow human laws and expectaions. As the authors say in their abstract "Phylogeny of the clade is interpreted from both (cladistic and, phenetic/stratigraphic) paradigms, and the interpretations are found to he inconsistent. Resolution of the phylogeny rests on the emphasis of one type of data over another. An interpretation of cladogenesis within the genus Neusticosaurus entails rejection of suggestive phenetic and stratigraphic data, whereas an anagenetic interpretation entails reversal of autapomorphies in ancestral taxa. Anagenesis is deemed to he the more probable interpretation, based on the strength of the stratigraphic and phenetic data relative to the character data. Implications of the test case results for phylogeny reconstruction in general are discussed, ending with a call for pluralism in approach" (O'Keefe & Sander 1999). The anagenetic hypothesis in this case is supported also by long bone microstructure, according to which Serpianosaurus has the more primitive, longitudinally orientated, vascularisation pattern, which is replaced by the more advanced, radially orientated, pattern of Neusticosaurus edwardsii (Hugi 2011)
For this reason, in the dendrograms at the top of the relevant pages here, instead of the usual branching pattern, the two general are connected by a single line (as Serpianosaurus--Neusticosaurus, and on this page all three species of Neusticosaurus are included. Strictly speaking of course the distinction between the two genera is quite arbitrary in any case, so Serpianosaurus could just as easily be included under Neusticosaurus. MAK111101
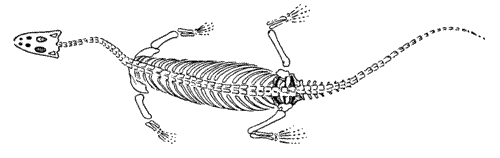 Neusticosaurus:
Seeley 1882.
N. ("Simosaurus") pusillus Fraas 1881 (includes N. "pygmaeus"
Fraas 1896 [5]); N. ("Pachypleurus")
edwardsii Cornalia 1854; N. peyeri Sander 1988. N.
("Psilotrachelosaurus") toeplitschi Nopcsa, 1929 is questionable.
N.
("Pachypleurosaurus") "staubi" Kuhn-Schnyder 1959 is a nomen dubium,
not diagnostic to the species level, the material of which belongs to either N.
peyeri or N. pusillus [CG85] [S89].
Neusticosaurus:
Seeley 1882.
N. ("Simosaurus") pusillus Fraas 1881 (includes N. "pygmaeus"
Fraas 1896 [5]); N. ("Pachypleurus")
edwardsii Cornalia 1854; N. peyeri Sander 1988. N.
("Psilotrachelosaurus") toeplitschi Nopcsa, 1929 is questionable.
N.
("Pachypleurosaurus") "staubi" Kuhn-Schnyder 1959 is a nomen dubium,
not diagnostic to the species level, the material of which belongs to either N.
peyeri or N. pusillus [CG85] [S89].
Range: Middle Triassic (Ladinian) to Late Triassic (early Carnian) of Europe (Northern Italy, SW Germany & Switzerland) [CG85] [S89].
Phylogeny: Pachypleurosauridae ::: Serpianosaurus + *.
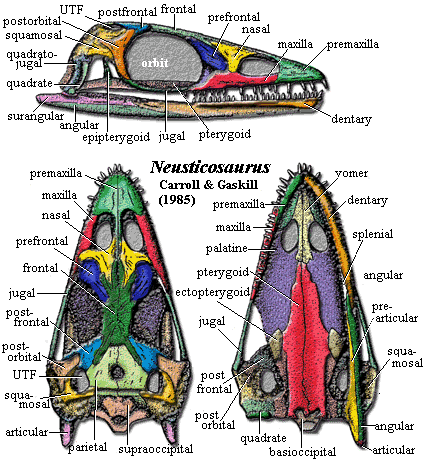 Characters:
dermal skull: skull development with strong negative allometry, very
small in large adults [CG85] [S89]; premaxillae broadly expanded across snout,
with posterior process separating nares, nasals [CG85]; sides of snout weakly
convex [R89]; lacrimal probably absent
in all species [S89]; maxilla with small or absent suborbital extension [CG85]
[S89]; nasals large, may be separated at midline by frontals & premaxillae
superficially (peyeri, pusillus) or completely (edwardsii) [CG85]
[R89] [S89]; nasal with curving process to
maxilla (in edwardsii only [S89]) [CG85]; nasal with anterior (to naris) and posterior processes [CG85];
nares formed by premaxilla, nasal and maxilla [S89]; frontals large, variably fused [CG85]
[S89]; frontals form dorsal orbit [S89]; large pineal [R95] [S89];
parietal without median ventral flange [CG85]; parietal lappet onto occipital
surface [CG85]; parietal (and squamosal [S89]) thickened medially to form transverse ridge dividing
the skull table from occiput [CG85]; UTF small, bordered by squamosal, parietal,
postfrontal & sometimes (narrowly) by postorbital [S89] [R89]; orbits large [CG85] [S89]; pre- & postfrontals
massive, suggesting bulging eyeball [CG85]; prefrontal forming anterior orbit
[S89]; sclerotic plates well-developed (14-20?) [CG85] [S89]; jugal extends anteriorly nearly to front of orbit. [CG85];
jugal forms ventral orbit [S89]; jugal slightly broadened
below postorbital [CG85]; postorbital skull short & narrow [CG85] [S89]; postorbital
triangular, covers squamosal anteriorly and almost bisects lateral exposure of
squamosal [CG85]; postorbital with deep medial process into postfrontal [S89];
postorbital ventral process articulates with end of jugal [S89]; squamosal
on lateral & posterior margin of small upper temporal fenestra [CG85] [R95];
squamosal also forms dorsal margin of lower temporal embayment [CG85]; squamosal
with lappet onto occipital surface [CG85]; squamosal with notch to receive
quadrate [CG85]; quadratojugal thin, but largely covering quadrate in lateral
view [CG85]; supraoccipital arched, extending below, but not sutured to,
parietals [CG85]; posttemporal fenestrae present [CG85]; exoccipitals very small
[CG85]; exoccipitals with single XII foramen and recessed for jugular foramen
[CG85];
Characters:
dermal skull: skull development with strong negative allometry, very
small in large adults [CG85] [S89]; premaxillae broadly expanded across snout,
with posterior process separating nares, nasals [CG85]; sides of snout weakly
convex [R89]; lacrimal probably absent
in all species [S89]; maxilla with small or absent suborbital extension [CG85]
[S89]; nasals large, may be separated at midline by frontals & premaxillae
superficially (peyeri, pusillus) or completely (edwardsii) [CG85]
[R89] [S89]; nasal with curving process to
maxilla (in edwardsii only [S89]) [CG85]; nasal with anterior (to naris) and posterior processes [CG85];
nares formed by premaxilla, nasal and maxilla [S89]; frontals large, variably fused [CG85]
[S89]; frontals form dorsal orbit [S89]; large pineal [R95] [S89];
parietal without median ventral flange [CG85]; parietal lappet onto occipital
surface [CG85]; parietal (and squamosal [S89]) thickened medially to form transverse ridge dividing
the skull table from occiput [CG85]; UTF small, bordered by squamosal, parietal,
postfrontal & sometimes (narrowly) by postorbital [S89] [R89]; orbits large [CG85] [S89]; pre- & postfrontals
massive, suggesting bulging eyeball [CG85]; prefrontal forming anterior orbit
[S89]; sclerotic plates well-developed (14-20?) [CG85] [S89]; jugal extends anteriorly nearly to front of orbit. [CG85];
jugal forms ventral orbit [S89]; jugal slightly broadened
below postorbital [CG85]; postorbital skull short & narrow [CG85] [S89]; postorbital
triangular, covers squamosal anteriorly and almost bisects lateral exposure of
squamosal [CG85]; postorbital with deep medial process into postfrontal [S89];
postorbital ventral process articulates with end of jugal [S89]; squamosal
on lateral & posterior margin of small upper temporal fenestra [CG85] [R95];
squamosal also forms dorsal margin of lower temporal embayment [CG85]; squamosal
with lappet onto occipital surface [CG85]; squamosal with notch to receive
quadrate [CG85]; quadratojugal thin, but largely covering quadrate in lateral
view [CG85]; supraoccipital arched, extending below, but not sutured to,
parietals [CG85]; posttemporal fenestrae present [CG85]; exoccipitals very small
[CG85]; exoccipitals with single XII foramen and recessed for jugular foramen
[CG85];
palate & braincase: quadrate not mobile [CG85]; quadrate
posterior margin embayed as otic notch [CG85] [S89]; otic capsule forming stout rod
extending toward top of cheek & supported by occipital flange of squamosal
[CG85]; stapes short cylinder [CG85]; basipterygoid articulation present, but
immobile [CG85]; epipterygoid thin, expanded at base [CG85]; sphenethmoid
probably ossified in part [CG85]; premaxilla forming broad shelf separated
posteriorly by vomers
[CG85] [S89]; premaxillary shelf participated in choana and meets maxillary
shelf [S89]; vomers narrow, meeting in weak suture [CG85]; choana large, round
[CG85]; maxilla borders choana, supports palatine laterally [S89]; palatine large, flat, thin [CG85]; pterygoids long, flat, solidly
attached to basioccipital [CG85]; pterygoid anterior process reaches vomer, but
not choana [S89]; pterygoid transverse flange essentially absent
[CG85]; pterygoid with parasagittal ridges medial to LTF (possibly pterygoideus
muscle) [CG85]; interpterygoid & suborbital
 vacuities closed by pterygoids [CG85]; ectopterygoid small, triangular, not
reaching cheek [CG85] (contra [S89]: pterygoid has thickened transverse
flange, ectopterygoid absent);
vacuities closed by pterygoids [CG85]; ectopterygoid small, triangular, not
reaching cheek [CG85] (contra [S89]: pterygoid has thickened transverse
flange, ectopterygoid absent);
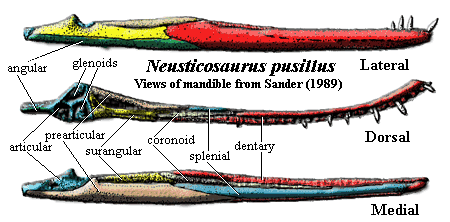
jaw: mandible thin, weakly attached at symphysis [CG85] (contra [S89], who points out that the dentaries are almost always found together); dentary weakly convex [R89]; dentary with large lateral exposure, bearing long groove for external adductor muscle attachment [CG85] [S89]; angular extends about 50% of jaw length [CG85]; surangular with dorsolateral recess for jaw muscles [CG85]; surangular extends into strong retroarticular process [S89]; articular forms ventral surface of retroarticular process, with transverse groove as articulating surface [CG85]; thecodont dentition [CG85] [S89]; teeth small, conical, with longitudinal striations [S89]; teeth flattened laterally & convex medially, with longitudinal striations (no keels) [S89]; 4-6 premaxillary teeth, with long roots [CG85]; anterior teeth slightly procumbent [CG85]; anterior maxillary teeth somewhat larger and caniniform [S89]; 17-25 total upper teeth [S89] 24-28 teeth on lower jaw [S89]; lower tooth row reaches well back of maxillary row [S89];
axial: axial elements variably pachyostosed, with considerable
inter- & intraspecific variation in degree and location [S89]; axis & atlas arches similar
to other cervicals [CG85]; 15-20 cervicals, 34-44 presacrals [S89]; centra, attachment area for arch covers almost entire
dorsal surface [CG85]; ribs on all presacrals [CG85]; neck long and at least laterally flexible [CG85];
cervical centra without adaptations for sharp bending
[CG85]; cervical centra
somewhat
 constricted medially, with longitudinal grooves [CG85];
cervical ribs double-headed [S89]; anterior cervicals
short (50% length of posterior cervicals), narrow, with shorter spines [CG85];
anterior cervical zygapophyses with larger (~20º) angle [S89]; posterior cervicals
with significant parapophyses [CG85]; anterior cervical ribs with distinct
anterior processes [S89]; cervical ribs larger posteriorly, sharply
pointed [CG85]; 19-24 dorsal vertebrae [S89]; anterior trunk probably stiffest
region of spine [S89]; trunk centra length increases over first 7, then constant
[CG85]; trunk centra swollen & "chubby" [CG85]; trunk zygapophyses broad, flat,
angled 5-10º [CG85]; trunk centra with extra median dorsal & ventral
articulations in some species [CG85] [S89] [1]; parapophyses
abruptly absent in trunk [CG85]; trunk transverse processes from arch only
except last two presacrals [CG85]; trunk spines tall, rhomboidal, slanted
posteriorly, with rugose dorsal surface (interspinous ligament) [CG85] ([S89]:
also in cervicals); trunk ribs
single-headed and attached to anterior portion of arch [CG85] [S89]; ribs pachyostosed in
adults [R95]; trunk ribs oriented laterally, abruptly changing to vertical, and
ending bluntly [CG85] [6]; 22-28 gastralia of three elements each, with 2 gastralia
to each corresponding vertebra [S89]; 3-4 sacrals [S89] [7]; sacrals with
extensive transverse processes, formed by both centra and arches [R89]; tail length quite variable [CG85];
40-60 caudals, 8-13 caudal ribs & 25-35 hemapophyses [S89];
caudal centra somewhat constricted medially, with longitudinal grooves [CG85];
caudal centra with arch attachment area quite narrow [CG85]; caudal zygapophyses
1-14 angled at ~20º [CG85]; large hemal arches from caudals 3/4 to 15, similar
to neural arches, then shrink rapidly [CG85]; hemal arches with rugose ventral
surface [CG85] [2]; last few
caudal ribs articulate only with centrum, other wise with both arch & centrum
[S89].
constricted medially, with longitudinal grooves [CG85];
cervical ribs double-headed [S89]; anterior cervicals
short (50% length of posterior cervicals), narrow, with shorter spines [CG85];
anterior cervical zygapophyses with larger (~20º) angle [S89]; posterior cervicals
with significant parapophyses [CG85]; anterior cervical ribs with distinct
anterior processes [S89]; cervical ribs larger posteriorly, sharply
pointed [CG85]; 19-24 dorsal vertebrae [S89]; anterior trunk probably stiffest
region of spine [S89]; trunk centra length increases over first 7, then constant
[CG85]; trunk centra swollen & "chubby" [CG85]; trunk zygapophyses broad, flat,
angled 5-10º [CG85]; trunk centra with extra median dorsal & ventral
articulations in some species [CG85] [S89] [1]; parapophyses
abruptly absent in trunk [CG85]; trunk transverse processes from arch only
except last two presacrals [CG85]; trunk spines tall, rhomboidal, slanted
posteriorly, with rugose dorsal surface (interspinous ligament) [CG85] ([S89]:
also in cervicals); trunk ribs
single-headed and attached to anterior portion of arch [CG85] [S89]; ribs pachyostosed in
adults [R95]; trunk ribs oriented laterally, abruptly changing to vertical, and
ending bluntly [CG85] [6]; 22-28 gastralia of three elements each, with 2 gastralia
to each corresponding vertebra [S89]; 3-4 sacrals [S89] [7]; sacrals with
extensive transverse processes, formed by both centra and arches [R89]; tail length quite variable [CG85];
40-60 caudals, 8-13 caudal ribs & 25-35 hemapophyses [S89];
caudal centra somewhat constricted medially, with longitudinal grooves [CG85];
caudal centra with arch attachment area quite narrow [CG85]; caudal zygapophyses
1-14 angled at ~20º [CG85]; large hemal arches from caudals 3/4 to 15, similar
to neural arches, then shrink rapidly [CG85]; hemal arches with rugose ventral
surface [CG85] [2]; last few
caudal ribs articulate only with centrum, other wise with both arch & centrum
[S89].
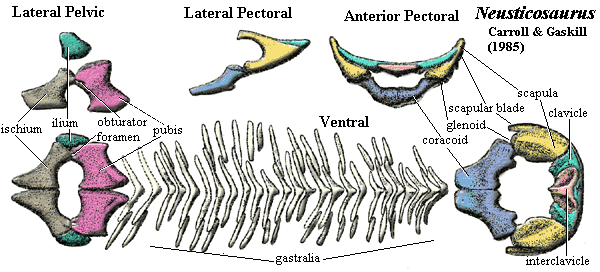
 appendicular:
[3] pectoral
girdle elements strongly bound, except coracoids [CG85]; pectoral fenestra
always present (but variable size) [S89]; interclavicle thin,
with short blade and rugose surface [CG85]; interclavicle with considerable
variation in shape and size; scapular blade short, rather narrow,
angled posteriorly at ~45º, with limited attachment surface [CG85] [S89]
[9];
suprascapular cartilage probably absent [CG85]; glenoid formed between scapula
& coracoid, but most cartilaginous [CG85]; coracoid flat, waisted [S89]; humerus with 50-60º
of tortion [CG85]; humerus length & shape with strong (presumably sexual)
dimorphism [S89]; humerus proximal end broadly oval, probably capped in
cartilage [CG85]; deltopectoral crest present, but poorly developed [CG85];
humerus distally flattened, with single distal articulation [CG85]; distinct
entepicondylar foramen [S89] [8]; ectepicondylar foramen restricted to a distal groove, lateral to ectepicondylar
ridge [CG85]; radius & ulna preserved as extending straight from end of humerus
[CG85]; ulna & radius about 50% length of femur, with ulna 80% length of
radius [CG85]; ulna broader than radius [R95]; radius tapers 50% distally &
extends beyond ulna [CG85]; forelimb with spatium interosseum
[R95]; olecranon absent [CG85]; only carpals are ulnare, intermedium and
(in edwardsii) a possible small lateral centrale [CG85] [S89]; ulnare circular [R95];
metacarpal I shorter than others, & III longest [R89]; metacarpals &
phalanges very slender [CG85]; manus phalangeal count 1-2,2-3,3,3-4,2 with
intraspecific variation in digits 3-5 [CG85]; pelvis mostly cartilaginous, with
considerable variation between species [CG85] [S89]; ilium small, triangular & constricted
dorsally, giving small sacral attachment area [CG85] [S89]; pubis & ischium
relatively similar, plate-like, flat, waisted, and without specialized medial
surface [CG85]; pubis separated from ischium by broad thyroid fenestra [CG85];
pubis with slit-like obturator foramen close to acetabulum [S89]; femur probably extended at 45º with lower limb directed posteriorly [CG85]
[4]; femur longer than humerus [R95]; femur slightly
curved [R95]; femur dorsoventrally flattened [CG85]; femur with expanded
proximal end, including trochanter major
(turtle term) possibly for iliofemoralis on posterior edge [CG85] [S89]; femur with "vaguely defined
intertrochanteric fossa" [CG85]; femur distally flattened [S89]; distal articulating surface only, with no
lateral development (as expected lower limb flexed much) [CG85]; no distinct
tibial & fibular condyles [CG85]; tibia & fibula short, stocky [CG85]; tibia
straight [R95]; medial edge of fibula strongly concave [CG85]; fibula distally
expanded "in the plane of the tarsals but is quite thin in the opposite
direction" [CG85]; fibula with distinct facets for astragalus & calcaneum
[CG85]; astragalus & calcaneum only ossified tarsals [CG85] [S89]; calcaneum circular
[CG85]; astragalus with crescentic indentation at articulation for tibia and
fibula [CG85] [S89]; metatarsals broad, and probably overlapping [CG85]; pes phalangeal count 1-2,2-3,3-4,4-5,3 with
intraspecific variation in digits 3-5 [CG85] [S89]; ungual phalanges wide, flattened &
bluntly rounded, possibly clawed in life (digits 1-3) [CG85] [S89].
appendicular:
[3] pectoral
girdle elements strongly bound, except coracoids [CG85]; pectoral fenestra
always present (but variable size) [S89]; interclavicle thin,
with short blade and rugose surface [CG85]; interclavicle with considerable
variation in shape and size; scapular blade short, rather narrow,
angled posteriorly at ~45º, with limited attachment surface [CG85] [S89]
[9];
suprascapular cartilage probably absent [CG85]; glenoid formed between scapula
& coracoid, but most cartilaginous [CG85]; coracoid flat, waisted [S89]; humerus with 50-60º
of tortion [CG85]; humerus length & shape with strong (presumably sexual)
dimorphism [S89]; humerus proximal end broadly oval, probably capped in
cartilage [CG85]; deltopectoral crest present, but poorly developed [CG85];
humerus distally flattened, with single distal articulation [CG85]; distinct
entepicondylar foramen [S89] [8]; ectepicondylar foramen restricted to a distal groove, lateral to ectepicondylar
ridge [CG85]; radius & ulna preserved as extending straight from end of humerus
[CG85]; ulna & radius about 50% length of femur, with ulna 80% length of
radius [CG85]; ulna broader than radius [R95]; radius tapers 50% distally &
extends beyond ulna [CG85]; forelimb with spatium interosseum
[R95]; olecranon absent [CG85]; only carpals are ulnare, intermedium and
(in edwardsii) a possible small lateral centrale [CG85] [S89]; ulnare circular [R95];
metacarpal I shorter than others, & III longest [R89]; metacarpals &
phalanges very slender [CG85]; manus phalangeal count 1-2,2-3,3,3-4,2 with
intraspecific variation in digits 3-5 [CG85]; pelvis mostly cartilaginous, with
considerable variation between species [CG85] [S89]; ilium small, triangular & constricted
dorsally, giving small sacral attachment area [CG85] [S89]; pubis & ischium
relatively similar, plate-like, flat, waisted, and without specialized medial
surface [CG85]; pubis separated from ischium by broad thyroid fenestra [CG85];
pubis with slit-like obturator foramen close to acetabulum [S89]; femur probably extended at 45º with lower limb directed posteriorly [CG85]
[4]; femur longer than humerus [R95]; femur slightly
curved [R95]; femur dorsoventrally flattened [CG85]; femur with expanded
proximal end, including trochanter major
(turtle term) possibly for iliofemoralis on posterior edge [CG85] [S89]; femur with "vaguely defined
intertrochanteric fossa" [CG85]; femur distally flattened [S89]; distal articulating surface only, with no
lateral development (as expected lower limb flexed much) [CG85]; no distinct
tibial & fibular condyles [CG85]; tibia & fibula short, stocky [CG85]; tibia
straight [R95]; medial edge of fibula strongly concave [CG85]; fibula distally
expanded "in the plane of the tarsals but is quite thin in the opposite
direction" [CG85]; fibula with distinct facets for astragalus & calcaneum
[CG85]; astragalus & calcaneum only ossified tarsals [CG85] [S89]; calcaneum circular
[CG85]; astragalus with crescentic indentation at articulation for tibia and
fibula [CG85] [S89]; metatarsals broad, and probably overlapping [CG85]; pes phalangeal count 1-2,2-3,3-4,4-5,3 with
intraspecific variation in digits 3-5 [CG85] [S89]; ungual phalanges wide, flattened &
bluntly rounded, possibly clawed in life (digits 1-3) [CG85] [S89].
N. pusillus Ladinian of Germany and Italy [R95]; short-lived but most widespread species [R95]; often associated with Lariosaurus. Significantly smaller than Serpianosaurus. [ORS99]
N. peyeri: closely related to N. pusillus and about the same size.
N. edwardsii: considerably larger than N. peyeri. This species is the source of a unique and probably unprecedented taxonomical conundrum: it has been contended that the holotype is not, in fact, a member of the species [ORS99]. In theory this can't happen.
Notes: [1] As [CG85] note, all these articulating structures "would presumably have made rotation of the vertebrae around the long axis nearly impossible, and might also have restricted lateral flexion of the trunk. Except where the vertebrae have become disarticulated from one another, the trunk region appears ramrod straight in these specimens. It appears to have formed an almost rigid structure in the living animal." [CG85: 369]. N. peyeri seems to have lacked these extra articulations, at least in the trunk. Sander's [S89] comments on the tail are ambiguous. He states [S89: 609] that accessory articulations are absent from the trunk; but, later on the same page, he says that "Accessory articulations are also found in [the proximal] portion of the tail" -- with no further elaboration.
[2] In essence there are two tail regions. The proximal region is deep, laterally flexible, strong and heavily muscled, with ribs and neural spines. The distal region is (impliedly) stiffer, rod-like and without muscles, ribs, or neural spines. The transition was relatively abrupt, occurring over 2-4 vertebrae.
[3] for a general description of the pectoral girdle, see Nothosauria.
[4] [CG85] note that this is similar to the marine iguana and crocodiles, both of which do not use the rear limb for propulsion. They add that the foot is, however, broad and might have served for steering.
[5] Important explanatory note: although N. pygmaeus Fraas 1896 is the same as N. pusillus Fraas 1881, the Fraas who named N. pusillus (Oscar Fraas), is not the same as the Fraas who named N. pygmaeus (Eberhard Fraas). On the other hand Oscar Fraas (1824-1897) is the same Fraas who named Eberhard Fraas (1862-1915) -- which he had a perfect right to do because Eberhard was Oscar's son, even though both men turned out to be members of the same species.
[6] Sander [S89] speculates that the dorsal ribs articulated with a cartilaginous sternum.
[7] The last dorsal rib and/or the first 1-2 caudal ribs are bent at odd angles. They look somewhat like sacrals and may have articulated with cartilaginous expansions of the pelvic bones [S89].
[8] The entepicondylar foramen is on the distal end in immature specimens, and its distance from the distal end of the humerus is a good proxy for maturity. However, it is also a sexually dimorphic character (see figure).
[9] Sander [S89: 612] states of N. peyeri that the scapular blade "extends back over the glenoid fossa, thereby limiting the dorsal excursion of the humerus." This would probably be true of all species. Carroll & Gaskill [CG85] also remark on the limited mobility of the forelimb in their study of N. edwardsii.
References: Carroll & Gaskill (1985) [CG85]; O'Keefe et al. (1999) [ORS99]; Rieppel (1989) [R89]; Rieppel (1995) [R95]; Sander (1989) [S89]. MAK991210, ATW061224, rev'd ATW080328.
Comment: Comparison of long bone microstructure in various species of Neusticosaurus and in Serpianosaurus indicate different habitats, longevities and ages at sexual maturity. The bone histological data regarding Serpianosaurus mirigiolensis and Neusticosaurus edwardsii also may indicate climate-dependant reproductive seasons similar to Recent lacertilian squamates. Hugi et al 2011). MAK111103
| Page Back | Unit Home | Page Top | Page Next |
ATW061231 revised ATW080328 checked ATW080127
Using this material. All material by ATW is public domain and may be freely used in any way (also any material jointly written by ATW and MAK). All material by MAK is licensed Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners