| Palaeos | Insectivora | |
| Vertebrates | Insectivora |
| Page Back | Unit Home | Unit Dendrogram | Unit References | Taxon Index | Page Next |
| Unit Back | Vertebrates Home | Vertebrate Dendrograms | Vertebrate References | Glossary | Unit Next |
|
Abbreviated Dendrogram
EUTHERIA | |--INSECTIVORA | `--+--Micropternodontidae | `--+--Erinaceomorpha | `--Soricomorpha | |--Tenrecoidea | `--+--Solenodontidae | `--+--Talpoidea | `--Soricoidea | |--Apternodontidae | `--Soricidae | |--Crocidurinae | `--Soricinae `--+--Chrysochloroidea `--AFROTHERIA |
Contents
Index |
The Insectivora have long been recognised as being amongst the most primitive of placental mammals, although precise taxonomy and phylogeny and their distinction from other early unspecialise dmammals was difficult because of their generalised nature. As a result they became something of a wastebasket taxon in which any small, unspecialised Cretaceous to Early Tertiary fossil mammals were included. Simpson 1945 in the first comprehensive evolutionary systematic taxonomy of all known living and extinct mammals, includes under the Order Insectivora five extant (Tenrecoidea, Chrysochloroidea, Erinaceoidea, Macroscelidoidea, and Soricoidea) and three fossil (Deltatheridioidea, Pantolestoidea, Mixodectoidea) superfamilies. Colbert 1969 pp.288-9, 484, lists four suborders, while acknowledging the unstatisfactoriness of this schema: the proteutherians or menotyphlans (a grab bag of early fossil types such as those mentioned by Simpson: leptictids, pantolestids, Zalambdalestes, etc,), macroscelidids (elephant shrews), dilambdodonts or lipotyphlans (hedgehogs, shrews, and moles), and the zalambdodonts (tenrecs and golden moles).
The last few decades have seen a radical revision of the Insectivora, with many fossil and recent taxa previously included here relocated elsewhere. This has been due in part because of new discoveries and studies of Createcous and early tertiary forms, as well as new methodologies such as cladistics and molecular phylogeny. The proteutheria are pretty much acknowleged to be an artifical group (or at best paraphyletic) while the zalambdodonts are based on homoplasies such as a molar witha single V-shaped ridge (ironically a clade consisting of Tenrecoidea + Chrysochloroidea would later be revived through molecular sequencing)
The first cladistic analysis of extant mammals by Novacek 1986 remved groups such as elephant shrews and tree shrews that had previously been included under the Menotyphla, whilst leptictids were shown to be stem insectivora. He proposed the following insectivore phylogeny and taxonomy
Superorder Insectivora
Order Leptictida
Order Lipotyphla
Suborder Erinaceomorpha
Suborder Soricomorpha
Superfamily Tenrecoidea
Superfamily Soricoidea
While in their definitive review, McKenna & Bell (1997) renamed the Insectivora as Grandorder Lipotyphla, now consisting of three orders, Chrysochloridea, Erinaceomorpha, and Soricomorpha (although this seems to be an instance of taxonomic inflation).
A much more radical change resulted from the abaondonment of both linnaean evolutionary systematics and morphology-based cladistics. Here the situation is very different to reptile phylogeny, which involcves a large number of extinct taxa for which molecular sequencing is obviously not possible. In reptile phylogeny, the evolutionary systematic model of Romer et al was replaced by cladistic morphological analysis of Gauthier and co-workers, which strongly emphasised skeletal characters of fossil groups. But since most major mammal clades are still extant (apart from the Meridungulates and various Paleocene and Eocene forms) phylogenetic studies, molecular phylogeny was the new paradigm of choice, and indeed this developed with mammals at the sam etime cladistics did with reptiles.
The problem here is the huge incongruity between morphology-based and molecular-based phylogenies. Due to very different molecular sequences between the Eurasian and African insectivores, the two morphologically very similar groups has been divided among two phylogenetically quite unrelated groups of mammals, resulting in a polyphyletic insectivora, divided between the two major clades of Laurasiatheria and Afrotheria.
As molecular phylogeny has grown in popularity over the last decade or so, and a strong consensus has emerged among molecular studies, there has been a rejection of many previously well regarded monophyletic clades, not only among mammals, but reptiles and other groups as well. The present author gets the impression, from browsing online forums and current literature, that, all else being equal, and in any conflict between molocules and morphology, it is almost always assumed, that molecular phylogeny has the more reliable phylogenetic signal, and should be used in prefernce to a synapomorphy morphology-based approach.
We are not proposing that molecular phylogeny is worse than a cladistic synapomorphy based approach, but, like all methodologies, it can be highly problematic. Few seem to consider how radical the gross physiological homoplasies, reversals and so on would have to be to make this non-morphological phylogeny work. As to whether stratophenetic evolutionary systematics, cladistic morphological analysis, molecular sequencing, morphology + molocules total evidence approach, stratocladistics, or some combination of any or all of the above, is the best way to tease out an understanding of the subtle and complex path of the evolution of life on Earth, we leave to the reader to decide. As for the insectivora, we have adopted something of a compromise. The golden moles have been moved to the Afrotheria, as per molecular sequencing and their basal position in the standard cladistic insectivore tree, but their geographic compatriots the tenrecs have been retained in the Insectivora, as they are morphologically typical insectovores. MAK120320
 Insectivora (= Lipotyphla) : Batodon, shrews, moles, hedgehogs, tenrecs. etc.
Insectivora (= Lipotyphla) : Batodon, shrews, moles, hedgehogs, tenrecs. etc.
Range: originally considered from the Late Cretaceous [N99]; here revised to Early Paleocene (Wible et al 2007).
Phylogeny:
Morphology: Epitheria :: ((Anagalida + Archonta)
+ (Ferae + Ungulatomorpha)) + *: Chrysochloroidea + (Micropternodontidae + (Erinaceomorpha + Soricomorpha)).
Molecular: Laurasiatheria : (Ferungulata + Chiroptera) + * : Solenodontidae + Talpoidea + (Erinaceomorpha + Soricoidea) (polyphyletic, Chrysochloroidea and Tenrecoidea included in the Afrotheria)
Wible et al 2009: Eutheria ::: Laurasiatheria + (Euarchontaglires : Archonta + (Afrotheria + Xenarthra) + *) We are mostly following Wible et al 2009 except that the Xenarthra are here considered more basal MAK120319
Characters: Skull low and triangular [N99]; rostrum long and narrow
[N99]; rostrum mobile [MN93];
maxilla widely expanded [$C88]; maxilla with substantial orbital wing
extending into the middle of the orbit [MN93] [5] [A+02$?];
maxilla broadly contacting frontal in facial region [MN93]; lacrimal facial wing absent [$C88]
or much reduced [MN93];
orbits fairly small & many fossorial forms effectively blind
[V+00]; unique skull pattern from small orbit & large nasal capsules [$C88];
infraorbital canal generally short [MN93]; no postorbital bar [N99]; jugal reduced or absent [$C88] [MN93] (not
synapomorphic) [N99] [V+00];
zygomatic arch usually 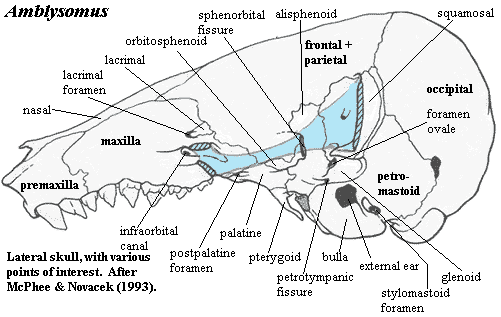 incomplete [$C88] [N99] [V+00]; post-glenoid foramen for
external jugular vein present [$C88]; occipital condyle, "ventral eminence" with sigmoid anterior border [MN93]; pinnae
usually small or absent [N99]; mastoid large, ossified and incorporates
tympanohyal [MN93]; large pyriform
fenestra between otic capsule and basisphenoid [$C88]; tympanic
annular [V+00]; auditory bulla frequently absent [C88] [V+00]; entotympanic frequently
absent [V+00]; tympanic cavity often covered by processes from adjoining bones
[V+00]; petrotympanic fissure present [MN93]; petrosal with distinct crest on
medial promontorium
[MN93]; alisphenoid surrounds fenestra
ovalis [V+00]; bony dorsum
sellae absent [$C88]; alisphenoid canal present [A+01$?]; large olfactory lobes mostly interorbital [V+00];
orbitosphenoid anterior to braincase (!?) [$C88] [V+00]; orbitosphenoid
large [MN93]; brain small & cerebral
hemispheres do not expand back over cerebellum [C88] [N99]; cerebrum smooth [C88]
[N99] [V+00]; palatine
reduced [$C88]; deciduous teeth shed early & rarely functional [N99];
all teeth rooted [N99]; dental formula typically 3/3, 1/1, 4/4, 3-4/3-4 [N99]; teeth with sharp cusps [V+00]; incisors large and
specialized [V+00]; I1 larger than other anterior teeth [A+02$?]; canines reduced [V+00]; molars often zalambdodont or dilambdodont; limbs unspecialized and never
saltatorial [V+00]; stance plantigrade, with 5/5 digits [C88] [N99]; clavicle
present (except in the tenrec Potamogale) [N99]; digits I&V not
opposable [C88] [N99]; pubic symphysis reduced (?) [$C88] [MN93$]
[A+02$?] [4];
tibia with lateral flange (obturator ridge) joining lesser & greater
trochanters [A+02$?]; tibia &
fibula frequently fused distally [N99]; lack of differentiated hindgut, with cecum absent,
large intestine absent, and short intestinal tract [C88] [MN93$] [V+00] [3];
testes internal [C88] [N99]; some groups with
cloaca [N99] [V+00]; hemochorial placenta [MN93]; pelage
of short, close-set fur, with fur not polymorphic, but may include spines [N99];
pelage with guard
hairs only; high metabolic rate; nocturnal (except some shrews) [N99]; territorial, but little social
structure.
incomplete [$C88] [N99] [V+00]; post-glenoid foramen for
external jugular vein present [$C88]; occipital condyle, "ventral eminence" with sigmoid anterior border [MN93]; pinnae
usually small or absent [N99]; mastoid large, ossified and incorporates
tympanohyal [MN93]; large pyriform
fenestra between otic capsule and basisphenoid [$C88]; tympanic
annular [V+00]; auditory bulla frequently absent [C88] [V+00]; entotympanic frequently
absent [V+00]; tympanic cavity often covered by processes from adjoining bones
[V+00]; petrotympanic fissure present [MN93]; petrosal with distinct crest on
medial promontorium
[MN93]; alisphenoid surrounds fenestra
ovalis [V+00]; bony dorsum
sellae absent [$C88]; alisphenoid canal present [A+01$?]; large olfactory lobes mostly interorbital [V+00];
orbitosphenoid anterior to braincase (!?) [$C88] [V+00]; orbitosphenoid
large [MN93]; brain small & cerebral
hemispheres do not expand back over cerebellum [C88] [N99]; cerebrum smooth [C88]
[N99] [V+00]; palatine
reduced [$C88]; deciduous teeth shed early & rarely functional [N99];
all teeth rooted [N99]; dental formula typically 3/3, 1/1, 4/4, 3-4/3-4 [N99]; teeth with sharp cusps [V+00]; incisors large and
specialized [V+00]; I1 larger than other anterior teeth [A+02$?]; canines reduced [V+00]; molars often zalambdodont or dilambdodont; limbs unspecialized and never
saltatorial [V+00]; stance plantigrade, with 5/5 digits [C88] [N99]; clavicle
present (except in the tenrec Potamogale) [N99]; digits I&V not
opposable [C88] [N99]; pubic symphysis reduced (?) [$C88] [MN93$]
[A+02$?] [4];
tibia with lateral flange (obturator ridge) joining lesser & greater
trochanters [A+02$?]; tibia &
fibula frequently fused distally [N99]; lack of differentiated hindgut, with cecum absent,
large intestine absent, and short intestinal tract [C88] [MN93$] [V+00] [3];
testes internal [C88] [N99]; some groups with
cloaca [N99] [V+00]; hemochorial placenta [MN93]; pelage
of short, close-set fur, with fur not polymorphic, but may include spines [N99];
pelage with guard
hairs only; high metabolic rate; nocturnal (except some shrews) [N99]; territorial, but little social
structure.
 Note: [1] were it not for the Insectivora, the
higher mammals would fall on a rather neat rodent - primate spectrum plus the ungulates. This group, along with perhaps the Carnivora, are what are what
reduce mammalian taxonomy to total chaos. [2] The study by [A+02] did not
include Chrysochloroidea. Hence, the synapomorphies of Insectivora may
apply to a more restrictive group. Accordingly, they are all labeled as
questionable. [3] Unfortunately, as recognized by [MN93], this type
of adaptation is also found in, e.g., insectivorous bats and
pangolins. [4] although this adaptation is found in all fossorial
mammals, the same adaptation is found in all Insectivora, whether
fossorial or not. In non-fossorial species the symphysis is secondarily re-created by juxtaposed pre-acetabular surfaces. Further, the
usual symphysis exists embryonically and is lost, or replaced by the unique
pre-acetabular contact [MN93]. [5] a possible synapomorphy,
according to [MN93]. A problem is posed by the Chrysochloroidea, who have
a rather unique orbital mosaic, and a possible reversal in the tenrec Potamogale.
However, these may be further derived states.
Note: [1] were it not for the Insectivora, the
higher mammals would fall on a rather neat rodent - primate spectrum plus the ungulates. This group, along with perhaps the Carnivora, are what are what
reduce mammalian taxonomy to total chaos. [2] The study by [A+02] did not
include Chrysochloroidea. Hence, the synapomorphies of Insectivora may
apply to a more restrictive group. Accordingly, they are all labeled as
questionable. [3] Unfortunately, as recognized by [MN93], this type
of adaptation is also found in, e.g., insectivorous bats and
pangolins. [4] although this adaptation is found in all fossorial
mammals, the same adaptation is found in all Insectivora, whether
fossorial or not. In non-fossorial species the symphysis is secondarily re-created by juxtaposed pre-acetabular surfaces. Further, the
usual symphysis exists embryonically and is lost, or replaced by the unique
pre-acetabular contact [MN93]. [5] a possible synapomorphy,
according to [MN93]. A problem is posed by the Chrysochloroidea, who have
a rather unique orbital mosaic, and a possible reversal in the tenrec Potamogale.
However, these may be further derived states.
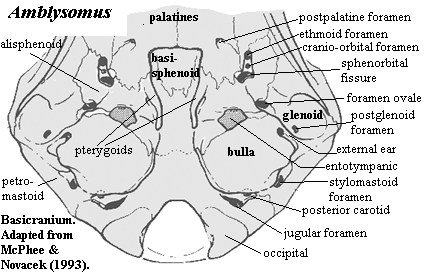
Image: Upper Right: Erinaceus from the Mammalian Crania Picture Archive of the Department of Anatomy, Tokyo University School of Medicine. Left & Lower Right: the Chrysochloroid (golden mole) Amblysomus hottentotus modified from [MN93].
Links: The Shrew (ist's) Site; Insectivora; IUCN/SSC - ITSES: Insectivore Specialist Group; Order INSECTIVORA; Order Insectivora; Mammalia: Insectivora; Entrez-PubMed; Entrez-PubMed; Order Insectivora; Insectivora; APUS.RU | Speesok Veedov | Mlekopeeta ... (Russian); Google Directory - Science >: Biology >: Flora and Fauna >: Animalia ...; Mammalogy Image Archives- Insectivora; Order Insectivora- EnchantedLearning.com; Insectivores - Order Insectivora; Insectivora; ???? (Chinese & English); Bibliography of Insectivora; ordo Insectivora; Order Insectivora- Solenodons; Untitled; Main Page.
References: Asher et al. (2002) [A+02]; Carroll (1988) [C88]; MacPhee & Novacek (1993) [MN93]; Nowak (1999) [N99]; Vaughan et al. (2000) [V+00]. ATW030720.
Micropternodontidae: Carnilestes, Micropternodus
Range: Early Paleocene to Early Miocene of China, Mongolia & North America
Phylogeny: Insectivora :: (Erinaceomorpha + Soricomorpha) + *.
Characters: infraorbital canal short [MN93]; maxillary root of zygomatic arch far back at M3 [A+01$?]; maxillary zygomatic rudiment reduced [A+01$?]; zygomatic arch incomplete, with jugal reduced or absent [A+01$?]; "no" zygomatic arch [MN93]; internal face of coronoid process deeply concave, but without pocket [A+01$?]; alveolar ridge on medial dentary running from m3 to condyle [A+01$?]; C1 with single root [A+01$?]; premolar series without diastema [A+01$?]; "semimolariform" P4 [MN93]; upper molars rectangular, with reduced stylar shelves; prominent metacone [A+02]; enlarged hypocones [MN93]; paraconids transversely oriented, crest-like; broad talonid basin [A+02].
Links: Micropternodontidae (Mikko's Phylogeny); Untitled Document (Carnilestes abstract).
References: Asher et al. (2002) [A+02]; MacPhee & Novacek (1993) [MN93]. ATW030731.
| Page Back | Unit Home | Page Top | Page Next |
checked ATW050623, revised MAK120319