| Palaeos | Aves | |
| Vertebrates | Ratites |
| Page Back | Unit Home | Unit Dendrogram | Unit References | Taxon Index | Page Next |
| Unit Back | Vertebrates Home | Vertebrate Dendrograms | Vertebrate References | Glossary | Unit Next |
|
Abbreviated Dendrogram
COELUROSAURIA | AVES |--Archaeornithes `--+--Confuciusornithidae `--Ornithothoraces |--Enantiornithes `--Ornithurae |--Hesperornithiformes `--Neornithes |--Paleognathae | |--Lithornithiformes | `--Ratites | |--Struthioniformes | `--+--Rheiformes | |--+--Aepyornithiformes | | `--+--Dinornithiformes | | `--Apterygiformes | `--Casuariformes `--Neoaves |--GALLOANSERAE `--GRUIMORPHA |
Contents
Index |
 About 30 years ago, Joel Cracraft
(1974) sketched out the central melodies of
ratite evolution. Ever since that time, he has been trying to get a very mixed chorus of avian paleontologists and ornithologists -- not to mention
ostriches, emu, moas,
cassowaries, and the like -- to learn the words, sing in
harmony, and keep to the rhythm. He has had some success. Everyone
has learned the words and music, the basic premises of Cracraft's
story. The ratites all descended from a common ancestor in the Late
Cretaceous. They evolved on the supercontinent Gondwana. When
Gondwana fragmented, these continents, like lost Israelite tribes, were scattered to the four
corners of the Earth -- or, more exactly, to four fragments of Gondwana --
and the ratite groups lost touch with each other. In Cracraft's scenario,
they subsequently diversified into their
present, separate forms. Then, perhaps, they will be reunited in a closing
production number
involving the said emu, moas, cassowaries etc., running at high speed across the
set, while assorted ornithologists and learnèd paleos dance a
stately pavane with elephant birds and kiwis at center stage. Finally, a
tinamous, lithornithid, or
similar awkwardly volant form, flies full tilt into the lighting booth with an
incandescent crash of exploding limelights. The set goes dark, and the curtain falls to thunderous
applause.
About 30 years ago, Joel Cracraft
(1974) sketched out the central melodies of
ratite evolution. Ever since that time, he has been trying to get a very mixed chorus of avian paleontologists and ornithologists -- not to mention
ostriches, emu, moas,
cassowaries, and the like -- to learn the words, sing in
harmony, and keep to the rhythm. He has had some success. Everyone
has learned the words and music, the basic premises of Cracraft's
story. The ratites all descended from a common ancestor in the Late
Cretaceous. They evolved on the supercontinent Gondwana. When
Gondwana fragmented, these continents, like lost Israelite tribes, were scattered to the four
corners of the Earth -- or, more exactly, to four fragments of Gondwana --
and the ratite groups lost touch with each other. In Cracraft's scenario,
they subsequently diversified into their
present, separate forms. Then, perhaps, they will be reunited in a closing
production number
involving the said emu, moas, cassowaries etc., running at high speed across the
set, while assorted ornithologists and learnèd paleos dance a
stately pavane with elephant birds and kiwis at center stage. Finally, a
tinamous, lithornithid, or
similar awkwardly volant form, flies full tilt into the lighting booth with an
incandescent crash of exploding limelights. The set goes dark, and the curtain falls to thunderous
applause.
It's the last bit that, perhaps not surprisingly, has caused some little difficulty. Cracraft's "vicariance" hypothesis, as such biogeographical theories are known, has an awful lot of moving parts. So, although Cracraft is probably correct, the participants have had considerable trouble choreographing their moves. Here we provide a review of the geography, phylogeny and timing of this 100 million year ("Ma") ballet, from which we have sternly excised (almost) all references to Swan Lake, L'Après-Midi d'Une Fauna, or The R(at)ite of Spring as unworthy of a serious discussion of avian phylogeny.
 Gondwana
was one of the first "supercontinents" on Earth. It is certainly
the oldest supercontinent known in any detail. It consisted of what are now the separate cratons of South
America, Africa, Madagascar, India, Australia (including New Guinea and New
Zealand), and East and West Antarctica (the last named originally formed as
two separate continents). Gondwana was first formed by a coalescence of
continents in the Southern Hemisphere after the break-up of Rodinia in
the Tonian Period, almost 900 Ma ago. We know quite about a bit about it
because it lasted a very long time. It remained the dominant landmass in
the Southern Hemisphere until the Oligocene,
only 30 Ma ago. At that time, the Drake Passage opened up between South
America and Antarctica, the last two pieces of Gondwana. The Drake Passage isolated Antarctica -- not only
geographically, but climatically. With the departure of South America, the
South Polar current could circle completely around Antarctica, without land
barriers. This continuous circling current acts like the continuous,
circling, moat of a castle, allowing no warmer water to enter. The prevailing
winds act in a similar fashion. The result is that Antarctica is not
simply the coldest place on Earth today. It is probably the coldest place on Earth
at any time since the aptly-named Cryogenian
Period, about 700 Ma ago.
Gondwana
was one of the first "supercontinents" on Earth. It is certainly
the oldest supercontinent known in any detail. It consisted of what are now the separate cratons of South
America, Africa, Madagascar, India, Australia (including New Guinea and New
Zealand), and East and West Antarctica (the last named originally formed as
two separate continents). Gondwana was first formed by a coalescence of
continents in the Southern Hemisphere after the break-up of Rodinia in
the Tonian Period, almost 900 Ma ago. We know quite about a bit about it
because it lasted a very long time. It remained the dominant landmass in
the Southern Hemisphere until the Oligocene,
only 30 Ma ago. At that time, the Drake Passage opened up between South
America and Antarctica, the last two pieces of Gondwana. The Drake Passage isolated Antarctica -- not only
geographically, but climatically. With the departure of South America, the
South Polar current could circle completely around Antarctica, without land
barriers. This continuous circling current acts like the continuous,
circling, moat of a castle, allowing no warmer water to enter. The prevailing
winds act in a similar fashion. The result is that Antarctica is not
simply the coldest place on Earth today. It is probably the coldest place on Earth
at any time since the aptly-named Cryogenian
Period, about 700 Ma ago.
In fact, ice-covered continents are not at all common in Earth's history, even at the poles. During the Cryogenian, the entire Earth may have been covered in ice, including the seas. However, since that time, substantial polar ice caps, much less true ice ages, have occurred only three times. It is possible that some cold spells have been so brief we have found no direct record of them However, polar ice caps lasting more than 1-2 Ma are known only from the Late Ordovician (short, but severe, ice age), the Carboniferous (mild, but very long period of ice at the South Pole), and during the last 25 Ma, particularly the last 3-4 Ma. What these cold facts tell us is that, up until the very end, Gondwana had a relatively mild climate. No mountains of ice were present to separate different groups of birds living in various parts of Gondwana, particularly around Antarctica. While Antarctica may not have been tropical paradise in the Late Cretaceous and Paleogene, it was an open corridor.
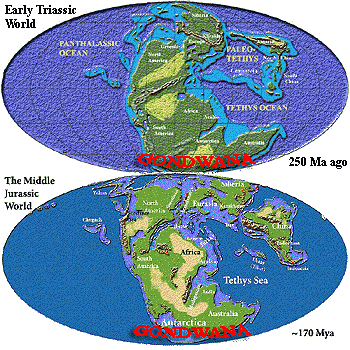
 Having
introduced our little family of continents, its time to show snapshots of the
kids. These are reduced copies of maps found elsewhere in Palaeos.
Much better maps are available elsewhere on the web in the usual places (e.g.
here and here).
We use our own only because they have no copyright, so we are free to abuse them
as we wish. As you can see -- if you have extremely good eyesight -- in
the Triassic, Gondwana was part of the world-continent of Pangea. That's Pangæa
if you live East of the Azores or in the Nineteenth Century. Antarctica
was the southern hub of Gondwana in those days. Africa and South America were a
single huge blob to the north and northeast, India and Madagascar formed a
peninsula to the north, and Greater Australia (including New Zealand and New
Guinea) was in the northeast. By the Middle Jurassic, some 80 Ma later,
all this continental coziness was beginning to fall apart. Eurasia and
North America, which had been living together since at least the Silurian,
decided that they really needed a little space, that space being the future
North Atlantic. South America and Africa have had an on-again-off-again
relationship for as long as anyone knows. In the Jurassic, they were
together, but both felt they'd had about as much of Antarctica as they could
stand and were both pulling away to the north. The little ones, India,
Madagascar, and Australia, were still solidly attached to mother Antarctica.
Having
introduced our little family of continents, its time to show snapshots of the
kids. These are reduced copies of maps found elsewhere in Palaeos.
Much better maps are available elsewhere on the web in the usual places (e.g.
here and here).
We use our own only because they have no copyright, so we are free to abuse them
as we wish. As you can see -- if you have extremely good eyesight -- in
the Triassic, Gondwana was part of the world-continent of Pangea. That's Pangæa
if you live East of the Azores or in the Nineteenth Century. Antarctica
was the southern hub of Gondwana in those days. Africa and South America were a
single huge blob to the north and northeast, India and Madagascar formed a
peninsula to the north, and Greater Australia (including New Zealand and New
Guinea) was in the northeast. By the Middle Jurassic, some 80 Ma later,
all this continental coziness was beginning to fall apart. Eurasia and
North America, which had been living together since at least the Silurian,
decided that they really needed a little space, that space being the future
North Atlantic. South America and Africa have had an on-again-off-again
relationship for as long as anyone knows. In the Jurassic, they were
together, but both felt they'd had about as much of Antarctica as they could
stand and were both pulling away to the north. The little ones, India,
Madagascar, and Australia, were still solidly attached to mother Antarctica.
However, the ratite story really concerns the last ages of Gondwana's long life. The next figure shows Earth in the Late Cretaceous, about 90 Mya. By this time, South America and Africa had broken up again. Europe and North America were still connected by shallow seas, but were only talking through their lawyers. Asia had become addicted to volcanoes and wasn't speaking to anyone. Back home with Mother Antarctica, Australia was still usually to be found at the apron strings, but India and Madagascar had gone off to live with Africa and didn't call home regularly. We'll talk more about that one later.
 In the meanwhile, we not only need more detailed maps, we need more detail
about map-making. Unlike maps of the present day, we have no
Cretaceous satellites to take photos. Ultimately, a paleomap rests on vast
amounts of data from paleomagnetism, sedimentary and bio-stratigraphy,
radiometric dating, biological and non-biological indicators of climate, stable
isotope and rare-element signatures, gravimetric data, a complex variety of
seismic (sonar-type) data, and so on. The science of paleogeography has
improved immeasurably over the last few decades. However, paleomap data
represents a sort of average over time. Since mapping requires that we
compare multiple data sets, and since date correlations between two kinds of
data are never precise, we can be very confident of the general result (because
the data sets overlap), but also confident that the map is only an average
(because we don't know exactly how far they overlap). The net
result is that, in addition to whatever minor noise might be generated by
measurement error, the data is generally only good to a precision of perhaps ±
4 Ma. Real coastlines change much more quickly, even without the movement
of continents. The port of Ostia, for example, was the main harbor for
Rome in the days of the Roman Republic. Ostia is now 3 km inland. If
that kept up for 10 Ma, Ostia would be about 15,000 km from the Mediterranean coast -- almost halfway around the world.
In the meanwhile, we not only need more detailed maps, we need more detail
about map-making. Unlike maps of the present day, we have no
Cretaceous satellites to take photos. Ultimately, a paleomap rests on vast
amounts of data from paleomagnetism, sedimentary and bio-stratigraphy,
radiometric dating, biological and non-biological indicators of climate, stable
isotope and rare-element signatures, gravimetric data, a complex variety of
seismic (sonar-type) data, and so on. The science of paleogeography has
improved immeasurably over the last few decades. However, paleomap data
represents a sort of average over time. Since mapping requires that we
compare multiple data sets, and since date correlations between two kinds of
data are never precise, we can be very confident of the general result (because
the data sets overlap), but also confident that the map is only an average
(because we don't know exactly how far they overlap). The net
result is that, in addition to whatever minor noise might be generated by
measurement error, the data is generally only good to a precision of perhaps ±
4 Ma. Real coastlines change much more quickly, even without the movement
of continents. The port of Ostia, for example, was the main harbor for
Rome in the days of the Roman Republic. Ostia is now 3 km inland. If
that kept up for 10 Ma, Ostia would be about 15,000 km from the Mediterranean coast -- almost halfway around the world.
Fortunately, that sort of thing does not continue for millions of years. The sedimentation of Ostia is an example of a short-term fluctuation. The problem is that, although such short-term fluctuations average out, no map can tell you what the coastlines were during any particular year of the Late Cretaceous. There are many sources of short-term variation. Most particularly, sea levels rise and fall more than you might expect. This was egregiously true during the Late Cretaceous. Sea levels were generally extremely high during this epoch. If you need to know why, look at Mesozoic: Climate. Since seas were high, they were covering a lot of gently-sloping continental shelf. Because the drop-off was very gradual, a small change in sea level might mean a huge change in the position of the shoreline. For these, and some other, related reasons, the Late Cretaceous is a particularly tricky time for map-makers trying to draw coastlines. As a very rough rule of thumb, and absent better information, we consider two adjacent continents separated by shallow, epicontinental seas (light blue) to be in biological contact, no matter how wide the gap. Conversely, two continents separated by deep ocean basin (dark blue), no matter how narrow the gap, are usually isolated. In the shallow sea case, distance does matter. The wider the gap, the slower or less likely it is to allow a species to move from one to the other. But that relates to our next topic, which is:
 But what exactly do we
mean by groups "losing touch" and why do we care? The "why
do we care" is, for once, the easier question. If we isolate a group
of organisms of the same species on an island with no internal barriers (i.e.
a freely interbreeding population) for a large number of generations, how many
species will we find descended from that original group? Ignoring the
possibility of extinction, the answer is likely to be 1. If the organisms
are freely interbreeding, then any mutations (purple antennae) are likely to be
lost (no one has purple antennae) or fixed in the gene pool (everybody
will have purple antennae) within a few generations, since the genes get mixed
quite thoroughly under these assumptions.
But what exactly do we
mean by groups "losing touch" and why do we care? The "why
do we care" is, for once, the easier question. If we isolate a group
of organisms of the same species on an island with no internal barriers (i.e.
a freely interbreeding population) for a large number of generations, how many
species will we find descended from that original group? Ignoring the
possibility of extinction, the answer is likely to be 1. If the organisms
are freely interbreeding, then any mutations (purple antennae) are likely to be
lost (no one has purple antennae) or fixed in the gene pool (everybody
will have purple antennae) within a few generations, since the genes get mixed
quite thoroughly under these assumptions.
Now, suppose we try the same experiment, but split the island into two halves with a big wall. What result when we knock down the wall after a few hundred generations? The most likely result is two separate species which are no longer capable of interbreeding. The mutations in the two populations will be different, and perhaps there will be some differences in selective pressures as well. In any case, the guys with purple antennae will have no interest in the girls with three eyes and vice-versa. Even if a pair forms, there's a good chance there will be no offspring, since the arrangement of genes on their respective chromosomes is likely to have changed, and even a slight change of that sort will make the cross either impossible or sterile.
This is a complex topic that we have grossly oversimplified. However, the basic point is correct. It is relatively hard to create two species out of a single, freely interbreeding population, whereas speciation is almost inevitable if you isolate a small group. I mean, after all, would you want your son going out with some little tart with three eyes?
So how much isolation is required? That depends on numerous factors. However, it is safe to say that the isolation has to be quite complete, although some cases are known in which gene flow was so slight that speciation occurred even in the presence of some contact. Waters & Roy (2004). It also helps if the isolation last for hundreds of generations, and if one or both populations are very small. If the gene pool is very small, random mutations are more likely to become fixed in the population, rather than diluted out and lost. Of course, there will also be fewer mutations in a small group, but the mathematics favor more rapid genetic drift in small populations. And, once a group all has purple antennae, its likely that they will be affected differently by the same environment. If nothing else, those with purple feelers are less likely to want to be out in freezing weather, to pick a trivial example. Thus, even if the original divergence is random, natural selection will kick in at some point, favoring mutations for cold-avoiding behaviors, hairy purple antennae, etc.
On the other hand, a small group is most vulnerable to genetic dilution. All it takes is a couple of studs with antennae to make it across the wall dividing our hypothetical island, before the three-eye trend has really caught on, and, very soon, you can forget about developing a separate population with three eyes. Thus the populations most likely to drift and create a new species are precisely those most vulnerable to dilution. Even a really tenuous genetic link can prevent the creation of a separate species if the original species is large, mobile, with long generation times -- like humans or ostriches, or even cats and dogs.
 Circular Reasoning
Circular ReasoningTo sum up the previous section, isolation means genetic isolation -- effectively zero transfer of genes for a number of generations proportional to the size of the isolated population. There are many exceptions to this general recipe for creating new species, one of which may be relevant to us.
In Southern California, south of the Central Valley, are two rather similar species of Ensatina, a salamander which lives in the mountains. Obviously all Ensatina derived from the same stock, but how? It turns out that, in the mountains north of the Central Valley lives a single species, intermediate between the two southern species. To make a long story short, these salamanders arrived in California from the north and worked their way around the Central Valley on the east and west sides. There is restricted, but continuous gene flow all the way around the Valley. However, by the time the salamanders had spread all the way round the Valley in both directions to meet near Los Angeles, the leading edges of eastern and western salamanders were sufficiently different that they could no longer interbreed. Actually, there are about seven species in the ring around the Central Valley, and more in a line following the Rockies into Canada. However, at any one place, we see only a continuous distribution of types, with any sufficiently motivated Ensatina able to mate with all of its (similarly motivated) neighbors of the opposite sex -- except in L.A. Of course, it being L.A., both species there are probably lying about their mating habits anyway. See Ensatina eschscholtzi for details and more examples.
Ratites (Ratitae): ostrich, emu, cassowary, kiwis.
Range: from the Paleocene.
Phylogeny: Paleognathae :: Tinamiformes + * : Struthioniformes + (Rheiformes + ((Aepyornithiformes + (Dinornithiformes + Apterygiformes)) + Casuariformes)).
Characters: Possibly polyphyletic assemblage of flightless paleognaths. Brain unusually small, but organized as in flying birds (e.g. large cerebellum); sternum generally reduced; keel absent; wing claws may be present in adults; tail may be elongated; "true" pygostyle absent(?); bone fusions weaker but generally as other neornithines; feather asymmetry essentially lost.
Links: Ratites Bibliography; Pour en savoir plus sur les ratites... (mostly ag links); Healthspa - Ratite Neuromuscular Diseases (veterinary information -- an interesting & unexplored possible phylogenetic angle); Birds - Everything About Ratites, Ostriches, Emus; Introduction to the Palaeognathae; The evolution of the palaeognathous birds; Read the Zoo- Ratites; Vorompatra Central- Ratites; ADW- Struthioniformes- Classification; Ratite;
Note: These sites are fairly random. There is a huge amount of agricultural information available for all living ratites because of their commercial value. Suggestions for additional scientific links would be very welcome. ATW021209.
| Page Back | Unit Home | Page Top | Page Next |
checked ATW050116