
| The Proterozoic Eon | ||
| Proterozoic | Proterozoic 2 |
| Archean | Proterozoic 1 | ||
| Phanerozoic | Timescale |
So, what were the earliest Metazoa? Apparently, they were the Vendobionta or Ediacaran organisms. They appear just before the end of the Marinoan Ice Age, more or less at the boundary between the Cryogenian and Ediacaran Periods. The oldest known representatives of the Vendobionta are probably the "Twitya disk" forms from the Twitya Formation in the McKenzie Mountains of northwestern Canada. Hofmann et al. (1990). These are pictured on the previous page. It remains unclear whether these are body fossils, or simply the holdfasts of something more elaborate. They include some very long-lasting "form taxa," (i.e. shapes without known animal shape-makers), and so the answer may be "holdfasts" (see also image 'a' below). However, that is unlikely to cover all of the disk-animals. In fact, Peterson et al. (2003) have expressed some doubt whether any of these structures -- even those plainly attached to other structures -- actually functioned as holdfasts.
Narbonne (2005) has reviewed the vendobionts relatively recently and has begun to connect some of the dots from the evolutionary data. His opinion is particularly helpful, because he has studied the extraordinary three-dimensional vendobiont fossils from Spaniard's Bay in Newfoundland (Canada) (image 'c' above). Narbonne follows Waggoner (e.g. Waggoner, 2003) in dividing the post-Twitya vendobionts into three assemblages: the Avalon, White Sea, and Nama.
 The Avalon Assemblage. The oldest, longest-lasting, and perhaps the best characterized assemblage is the Avalon. It appears between 600 and 560 Mya, and some elements of this fauna survived into the Cambrian. It is best known from several localities in Eastern Canada and from Charnwood Forest, England. However, elements of the fauna were quite cosmopolitan. They are found at virtually all depths and latitudes, limited only by the availability of wall-to-wall (bacterial) carpeting. Note that this kind of distribution is not quite what one might expect of a fauna expanding along some trend line of increasing ocean oxygenation -- but too little is known about the details of timing, ocean chemistry, and paleolocation to make much of it.
The Avalon Assemblage. The oldest, longest-lasting, and perhaps the best characterized assemblage is the Avalon. It appears between 600 and 560 Mya, and some elements of this fauna survived into the Cambrian. It is best known from several localities in Eastern Canada and from Charnwood Forest, England. However, elements of the fauna were quite cosmopolitan. They are found at virtually all depths and latitudes, limited only by the availability of wall-to-wall (bacterial) carpeting. Note that this kind of distribution is not quite what one might expect of a fauna expanding along some trend line of increasing ocean oxygenation -- but too little is known about the details of timing, ocean chemistry, and paleolocation to make much of it.
The members of this fauna fall into two morphological types: "medusoids" (disks) and the fern-like "fronds." Typical genera include Charnia, Ivesia, and Bradgatia. Narbonne calls the fronds "rangeomorphs" after the well-known Rangea [1]. In an important paper, Narbonne (2004) showed that the rangeomorphs appear to be built on a fractal architecture. That is each extension of the original structure gives rise to smaller structures which are "self-similar," i.e. have the same morphology as the parent structure. Close examination of the Spaniard's Bay rangeomorph in figure (c) above shows that each frond gives rise to "frondlets" of the same shape, which develop third level structures of the same shape, and so on. Contrast this pattern with some typical metazoan growth patterns, involving the addition of segments at the same scale along a single axis, or unsegmented and determinate growth.
 In addition to physical association and fractal development, the members of the Avalon Assemblage share a number of characters, including the complete absence of motility, the absence of any means of sediment clearance (as judged by mass-mortalities apparently caused by thin layers of sea-borne volcanic ash), a remarkable resistance to post-mortem compression, completely indeterminate growth, and a strong tendency to incorporate sediment centrally as a structural element. McMenamin (1998); Peterson et al. (2003). These features, including fractal development [2], are often found in the Fungi. Based on these similarities, Peterson's group speculates that many Avalon vendobionts were "analogous," or possibly related, to Fungi. We are less shy than Peterson et al. about the phylogenetic consequences. Whether the vendobionts are actually metazoans or not, it is hardly surprising to see organisms with a mosaic of metazoan and fungal characters a few tens of My before animals first become obvious in the fossil record. This is exactly what we ought to see at or near the base of Metazoa (toads > toadstools).
In addition to physical association and fractal development, the members of the Avalon Assemblage share a number of characters, including the complete absence of motility, the absence of any means of sediment clearance (as judged by mass-mortalities apparently caused by thin layers of sea-borne volcanic ash), a remarkable resistance to post-mortem compression, completely indeterminate growth, and a strong tendency to incorporate sediment centrally as a structural element. McMenamin (1998); Peterson et al. (2003). These features, including fractal development [2], are often found in the Fungi. Based on these similarities, Peterson's group speculates that many Avalon vendobionts were "analogous," or possibly related, to Fungi. We are less shy than Peterson et al. about the phylogenetic consequences. Whether the vendobionts are actually metazoans or not, it is hardly surprising to see organisms with a mosaic of metazoan and fungal characters a few tens of My before animals first become obvious in the fossil record. This is exactly what we ought to see at or near the base of Metazoa (toads > toadstools).
Two other structural observations from Narbonne's (2004) study of the Avalon fauna from Spaniard's Bay are worth mentioning. The first is that these vendobionts have more structural support than was supposed. It is hard to tell in most images, but Narbonne indicates that rangeomorphs were supported by bracing which formed an "organic skeleton" below the branching elements. The word "below" here is important. Although most reconstructions show rangeomorphs with a fern-like upright habit, a significant number of workers have expressed the view that rested on, or even buried in the sediment. See, e.g., Grazhdankin & Seilacher (2002). However Narbonne found (a) bracing which would be superfluous for a supine organism and (b) that both sides appeared identical in partially overturned specimens, again suggesting that both sides were exposed to the same environment. At least some of the frond animals therefore seem to have been erect.
Finally, Narbonne illustrates, but does not discuss, something we can only call the "corn cob" rangeomorph. This beast illustrates three structural themes of many vendobionts. First, as Narbonne notes, the "frondlets" which form the basic structural unit can bend, curl up, merge etc. and form many different shapes, including marginal rows of buttons. That marginal row looks disturbingly like Kimberella's marginal ornament, to our way of thinking. The second motif is the envelope which surrounded the corncob beast for at least 2/3rds of its circumference. We wonder how common this is. Many good images of vendobiont fossils suggest something of the sort. A few species (Ernietta) are acknowledged to have some such structure. McMenamin (1998). Could the mantle of Kimberella be derived from such an envelope?
 One thing is certain. The corncob animal cannot have been erect. At least some of the frond animals were therefore supine. However, it may also have been a reproductive part, embryo, or developmental resting stage. Vendobionts are never easy.
One thing is certain. The corncob animal cannot have been erect. At least some of the frond animals were therefore supine. However, it may also have been a reproductive part, embryo, or developmental resting stage. Vendobionts are never easy.
The White Sea Assemblage. The White Sea assemblage is found in sandy bottoms in temperate paleolatitudes during a relatively brief time span, from 556 to 550 Mya (Waggoner, 2003) and possibly as late as the base of the Cambrian (Narbonne, 2005). "Characteristic fossils of this assemblage include the bilaterally symmetrical forms, such as dickinsoniids, Kimberella, and Spriggina; the annulated concentric forms Kullingia and Ovatoscutum; and all but one of the triradially symmetrical discoid forms, such as Tribrachidium." Waggoner (2003: 107). "Worm burrows" suggest the presence of bilaterians (Narbonne, 2005), although trace fossils are notoriously difficult to interpret. This fauna is most diverse in relatively shallow waters (Narbonne, 2005).
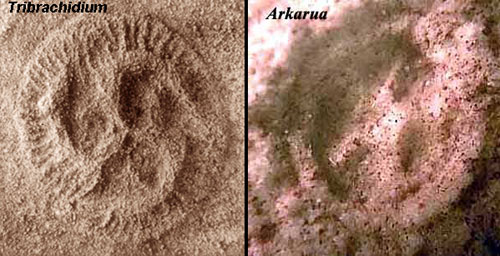
One of the seriously peculiar things about some vendobionts is their fundamental geometry. Two well-known examples are the triradial symmetry of Tribrachidium, and the pentaradial symmetry of Arkarua, both White Sea medusoids from Australia. Many vendobionts exhibit an alternating symmetry, with structures diverging from a central axis on alternate sides. In some cases, this is the result of flattening a fundamentally spiral pattern of growth. In others (particularly from the White Sea Assemblage), alternating growth seems to be the genuine body plan.
 Tribrachidium could conceivably be interpreted as an abbreviated spiral pattern. Some remains of Tribrachidium seem to show the three main arms at very different vertical levels. On the other hand, Pteridinium, which looks bilaterally symmetrical at first glance, seems to be based on a truly triradial plan, with two long branches forming the "segments" and one short branch forming a sort of spine. McMenamin (1998); Grazhdankin & Seilacher (2002).
Tribrachidium could conceivably be interpreted as an abbreviated spiral pattern. Some remains of Tribrachidium seem to show the three main arms at very different vertical levels. On the other hand, Pteridinium, which looks bilaterally symmetrical at first glance, seems to be based on a truly triradial plan, with two long branches forming the "segments" and one short branch forming a sort of spine. McMenamin (1998); Grazhdankin & Seilacher (2002).
The features of the White Sea communities which have attracted the most attention are (a) trace fossils strongly suggesting mobility; (b) some sort of segmentation (almost always of the alternating symmetry type); (c) a clear anteroposterior axis and (d) Kimberella, which has all of the above to one extent or another, as shown in the remarkable image from Fedonkin (2003).
Based on such characteristics, numerous workers have associated various vendobionts with specific taxa of living organisms, e.g. Kimberella with Mollusca, segmented forms with Anellida or Arthropoda generally, medusoids with Cnidaria or even Echinodermata. Others have been referred to as "soft-bodied trilobites." McMenamin (1998) (arguing that there is actually no close relationship). We will resist the temptation to use more muscular language here, and simply note that the evidence is not yet strong enough for any firm conclusions. After all, no one would suppose that rangeomorphs actually were ferns, or Ernietta a mushroom, based on basic morphology and lack of motility.
Perhaps, given the present strength of the scientific tide dragging the vendobionts into crown group animals (last common ancestor of all living animals and all of its descendants), it may be worth a few more contrarian comments to explain our doubts. The key characteristics are symmetry, metameric organization, and motility. As we have seen, vendobiont metamerism and symmetry are peculiar and often show a radial or alternating-side pattern. That pattern is more like a plant than an animal. And we have seen from Horodyskia and the annelidomorph algae that metameric (repeating) structures are not necessarily unique to crown group animals.
 Similarly, while vendobionts and animals both have large, motile forms, the vendobiont style of motility is peculiar. Fedonkin (2003) not only provides the image of Kimberella traces shown here, but also an image of similar traces radiating over 360° (or close to it -- the block is broken off in one sector). Whatever this preserves, it cannot be locomotion. From the same area of the White Sea, he reproduces Yorgia together with lighter imprints of the same organism, possibly the same individual. Together, they seem to show discontinuous movement, like a series of left footprints ending with the remains of a left foot. Quite possibly, this represents locomotion, but locomotion unlike any animal plausibly present in the Ediacaran. At the end of the day, there is good reason to suspect that the vendobionts are metazoans; but we have a hard time seeing them as crown group animals.
Similarly, while vendobionts and animals both have large, motile forms, the vendobiont style of motility is peculiar. Fedonkin (2003) not only provides the image of Kimberella traces shown here, but also an image of similar traces radiating over 360° (or close to it -- the block is broken off in one sector). Whatever this preserves, it cannot be locomotion. From the same area of the White Sea, he reproduces Yorgia together with lighter imprints of the same organism, possibly the same individual. Together, they seem to show discontinuous movement, like a series of left footprints ending with the remains of a left foot. Quite possibly, this represents locomotion, but locomotion unlike any animal plausibly present in the Ediacaran. At the end of the day, there is good reason to suspect that the vendobionts are metazoans; but we have a hard time seeing them as crown group animals.
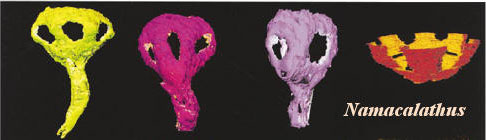
 The Nama Assemblage. The Nama Assemblage is somewhat younger than the White Sea communities. Its limits are approximately 549 to 542 Mya. That is, it overlaps the earliest Cambrian. Like the White Sea fauna, the Nama creatures are more abundant in shallow waters. Unlike the White Sea fauna, the Nama Assemblage is found at tropical paleolatitudes and in carbonate settings. Typical members are Namacalathus (image from Grotzinger et al., 2000) and Cloudina. These have been discussed elsewhere in our unfortunately chaotic and badly written essay on the origin of certain sponges. A recent addition with characters conforming to the model is Namapoikia. Wood et al. (2002).
The Nama Assemblage. The Nama Assemblage is somewhat younger than the White Sea communities. Its limits are approximately 549 to 542 Mya. That is, it overlaps the earliest Cambrian. Like the White Sea fauna, the Nama creatures are more abundant in shallow waters. Unlike the White Sea fauna, the Nama Assemblage is found at tropical paleolatitudes and in carbonate settings. Typical members are Namacalathus (image from Grotzinger et al., 2000) and Cloudina. These have been discussed elsewhere in our unfortunately chaotic and badly written essay on the origin of certain sponges. A recent addition with characters conforming to the model is Namapoikia. Wood et al. (2002).
The Nama Assemblage is more likely than the White Sea group to include the last common ancestor of demosponges and Demosthenes. If nothing else, members of this assemblage have calcified skeletons. In particular, various members have a mosaic of characters similar to well-known metazoans in or very close to crown group animals: inverted conical shapes, perforated exoskeletons, some double-walled regions, twisty "thrombolitic" internal structures, curved structural bracing, association with calcifying bacteria. These are characteristics found in archaeocyaths (probable sponges just basal to crown group animals) and some of the "small shelly fauna" both abundant in the Terreneuvian Epoch (earliest Cambrian). Interestingly, the Nama animals lack all of the putative animal characteristics of White Sea organisms. None are motile, bilaterally symmetrical, segmented, able to leap tall buildings in a single bound, etc.
We have, obviously, been unable to disguise our bias here, in spite of our earlier promises to the contrary. Having once again dynamited our own credibility, we will make a transparent attempt to salvage it by warning the reader, once again, not to take the argument too seriously. The current prevailing view is that Kimberella is a mollusk and that the inhabitants of the White Sea fauna were largely members of advanced animal clades, while the Nama assemblage consists of relatively uninteresting niche specialists.
It may be appropriate to close this section with a reference to the unnamed "Dengying form" of Xiao et al. (2005). The morphology of the living animal is unclear. It plainly doesn't fit well into any of the three conventional fauna described above. The overall plan is a sort of cross between a branching Avalon form and a"quilted" White Sea organism. However, it may have been weakly calcified, like a Nama creature, and some of the detailed cross-sections look vaguely stromatoporoid. If nothing else, the Dengying form should remind us that this part of phylospace is still mostly unexplored, and almost anything is still possible. ATW081218.