| Palaeos: | Sarcopterygii | |
| The Vertebrates | Tristichopteridae |
| Page Back | Unit Home | Unit Dendrogram | Unit References | Taxon Index | Page Next |
| Unit Back | Vertebrates Home | Vertebrate Dendrograms | Vertebrate References | Glossary | Unit Next |
|
Abbreviated Dendrogram
Teleostomi |--Neopterygii `--Sarcopterygii |--+--Onychodontiformes | `--Actinistia `--Rhipidistia |--Dipnomorpha `--Tetrapodomorpha |--Rhizodontiformes `--Osteolepiformes |--Tristichopteridae | |--Eusthenopteron | `--+--Cabonnichthys | `--Mandageria `--Elpistostegalia |--Panderichthys `--Tetrapoda |
Contents
Overview |
Let us admit what cannot be hidden. This is, quite obviously, a catch-all section for features that didn't fit well into other, better thought-out sections of this extended discussion. Nonetheless, there are features of interest here, albeit matters that are not easy to pigeon-hole into thematic boxes.
Most of the pertinent features of the palate have been discussed in our coverage of the braincase. The rhipidistian palate is a rather conservative structure, not that greatly different from the familiar tetrapod palate. Refer to our usual image of Eusthenopteron. The palate is dominated by a large pterygoid, a dermal bone which follows the ancient outline of the palatoquadrate, twisting to become a vertically oriented wall posteriorly, leaving a lateral subtemporal fossa for the descent of the jaw adductors. Anteriorly, it is bounded laterally by the inner tooth-bearing elements, the palatine and ectopterygoid, and medially by the long, denticulated palatal exposure of the parasphenoid. In osteolepiforms, the basicranial fossa leaves a gap in the middle of the palate which is roofed over by parotic (or arcual) plates. At the anterior end of the palate are found the palatal exposure of the premaxilla, the vomers, and often an anterior palatal recess between them.
Primitively, the vomers were variable in shape and position and seem to have wandered about fairly freely. In tetrapodomorphs, median support is critical, and the vomers always contact each other on the midline. As discussed previously, the vomers further develop a posterior process which tightly clasps the parasphenoid in tristichopterids. In derived tristichopterids, this relationship is further elaborated with "accessory vomers" which extend the contact even further posteriorly. By contrast, elpistostegalians have relatively minimal posterior vomerine processes and lack accessory vomers, perhaps for the reasons previously discussed.
There is a tendency for the vomers to move posteriorly. This is unsurprising, as it reduces the torque penalty. However, this displacement leaves both the choanae and the anterior palatal recess as holes in the palate adjacent to the margin, potentially weakening the jaw. Consequently, a number of lineages developed a median premaxillary process to bridge the anterior recess (e.g. Megalichthyidae), or extended a ventral median process from the braincase to accomplish the same task (Mandageria). Again as we might expect, Panderichthys accepted the marginal weakness for the sake of reducing torque. In fact, it maintained a wide anterior palatal recess and elongate choanae! The parasphenoid is always long in rhipidistians. In many forms it is also more or less stout and heavy, e.g., Youngolepis, Osteolepididae, and Cabonnichthys. However, consistent with our general take on Panderichthys, its parasphenoid is very thin and light.
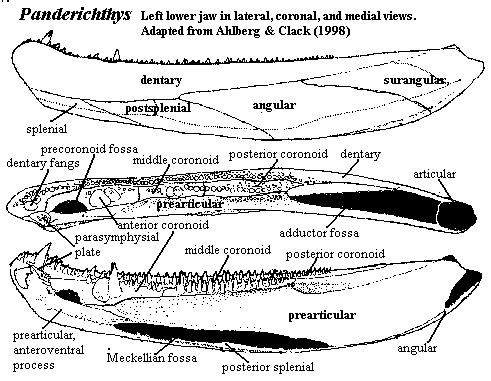 The
Lower Jaw
The
Lower JawAs Ahlberg & Clack (1998) showed us a few years ago, it is impossible to find any consistent phylogenetic signal in the structure of the lower jaw, at least in this part of phylospace. As the structure does not differ a great deal within the Tetrapodomorpha, we will not discuss it in any detail, save to point out that it remains essentially a hollow tube around the Meckelian cartilage, which is variably ossified.
One possible trend, of uncertain significance, is the reduction of the parasymphysial plate, which becomes quite small independently in both the Rhizodontiformes and the Tristichopteridae + Elpistostegalia. Among the elpistostegalians, there may be a general tightening up of the jaw with the reduction or elimination of the deep fossae before and between the coronoid series. By the level of Livoniana, the last and most persistent of these, the precoronoid fossa, has been completely closed. Presumably, this set the stage for the development of the comparatively rod-like rami of the amniote jaw. Another tendency is for the articular surface of the articular to face somewhat more dorsally, rather than being oriented posteriorly. Clack (2002). However, the overall structure of the lower jaw seems to have been tightly constrained, and subject only to the most gradual adjustment.
 The
Gill Skeleton
The
Gill SkeletonOne portion of the lower jaw which we conspicuously avoided discussing in the last paragraphs is the ventral component, consisting of a large median gular plate, lateral gulars, submandibulars, anterior extensions of the basibranchium, and possibly an anteriorly extended interclavicle. Unfortunately, these elements are often incompletely ossified, are frequently lost or damaged, are difficult to reconstruct, and are not often studied. Accordingly, there is almost nothing we can say with confidence.
Much the same is true of the branchial skeleton. However, two points can be made with some certainty. First, the hyomandibula and its articulations are relatively well known. It is double-headed in all rhipidistian groups, and articulates with the braincase at two points along the posterior margin of the lateral commisure. Primitively, the ventral articulation is directly below the dorsal articulation and both are located above the anterior part of the vestibular fontanelle. In derived tristichopterids and, independently, in elpistostegalians, the articulation moved posteriorly relative to the fontanelle and came to be located above the posterior portion of the fontanelle. Unfortunately, it is impossible to ascribe any particular meaning to this trend, since the rest of the hyoid arch is not known well enough in a sufficient number of genera to state whether this had some mechanical significance for the jaw, or merely reflected a change in the proportions of the otoccipital braincase.
As Johanson & Ahlberg (1997) have shown, the branchial skeleton has actually altered less than was traditionally thought. These authors not only analyzed the exceptionally well-preserved gill arches of Mandageria, but reinterpreted previous work on Eusthenopteron and other rhipidistians. They conclude that all rhipidistians have essentially the same arrangement. There are always three hypobranchials ("HB" in the figure) articulating with the basibranchium. There is always a fourth branchial arch (never a fifth) and its hypobranchial articulates on hypobranchial 3. Thus, the proportions may change from one taxon to another, but the structure is completely consistent in all rhipidistian groups. ATW031102.
GO TO PART 5. The Appendicular Skeleton
The tristichopterids were the most diverse and successful of the tetrapodomorph fishes throughout the Late Devonian, but were extinct by the end of the Famennian. Given the very poor fossil record of the early Mississippian, they may have continued briefly into the Carboniferous, but no further. The reasons for their disappearance are obscure. Other rhipidistian groups did well. Dipnoiforms, osteolepidids and megalichthyids all sailed past the end of the Famennian. Rhizodonts and tetrapods thrived. Perhaps a clue may be found in New South Wales, beside the road from Canowindra to Gooloogong. The huge Frasnian death assemblage [1] found there has a peculiar composition. It consists largely of antiarch placoderms. But, among the thousands of remains are also arthrodires, a canowindrid, and two different tristichopterids. So far as we are aware, the following taxa are very rare or absent: dipnoiforms (2 Soederberghia), osteolepidids (absent), megalichthyids (absent), rhizodonts (three of the aberrant Gooloogongia) and elpistostegalians / tetrapods (absent). So the fishes that died at Canowindra are mostly those that died and became extinct everywhere else within a few million years. The correspondence is hardly exact, and possibly this is all just a coincidence. Then again, perhaps not.
What is exceedingly odd about the disappearance of tristichopterids is that they are very similar to elpistostegalians, the immediate ancestors of tetrapods. Almost all of the characters of tristichopterids are shared by elpistostegalians, whether by inheritance or convergence. We have discussed what appears to be one distinction: the tendency to reinforce the cranial arch anteriorly, rather than lighten the rostrum as in Panderichthys. This may be the only significant qualitative distinction.
However, with that said, there are some differences of degree. Tristichopterids tend to be bigger fish than the known elpistostegalians. The body form is generally triangular in cross section. That is, tristichopterids have a flattened ventral surface, but are not dorsoventrally flattened overall. The skull may be flattened, but again does not reach the degree of dorsoventral compression which the elpistostegalians were believed to have achieved routinely. The tristichopterids also possessed a small postspiracular bone, a synapomorphy of the group, that may have closed up the spiracular (or otic) notch. In elpistostegalians, the absence of this bone, combined with the flatness of the head, may have caused this notch to flare very widely. In the axial skeleton, the pleurocentra are small, yet not as understated as in Panderichthys. Eusthenopteron, at least, has tetrapod-like ribs, but the shaft never develops ventrally. Eusthenopteron has "incipient" zygapophyses -- more than Panderichthys had -- but both features are missing in most derived genera.
These differences are rather unsatisfactory, and there are other considerations. We will take these up in succeeding sections. ATW031103.
Image: Eusthenopteron fossil from PAST LIVES- CHRONICLES OF CANADIAN PALEONTOLOGY - Chapter 22.
[1] Latest Frasnian or perhaps early Famennian. This assemblage and the circumstances of its discovery are discussed elsewhere.
Tristichopteridae: Devonosteus, Cabonnichthys, Eusthenodon, Eusthenopteron, Hyneria [2], Jarvikina, Litoptychius, Mandageria, Marsdenichthys, Notorhizodon, Platycephalichthys, Spodichthys?, Tristichopterus.
Range: Givetian to Famennian [CA96] of Europe, North America, Greenland, Russia, Antarctica & Australia.
Phylogeny: Osteolepiformes:::: Elpistostegalia + *: Eusthenopteron + (Cabonnichthys + Mandageria).
 Characters:
some very large (several meters); lateral rostral does not contact prefrontal
[AJ97] [1]; parietals usually long & narrow [AJ97]; postspiracular bones present [JA97] [AJ97];
primitively, lateral extrascapulars are widely separated [JA97]; prefrontal
contacts anterior tectal [JA97]; crista
parotica anteroposteriorly oriented [JA97]; ethmosphenoid cranial
segment elongate [JA97]; vomer posterior processes clasping sides of
parasphenoid [AJ97]; parasymphysial plate small, not contacting coronoid
[JA97]; basal scutes on
unpaired and unpaired fins [AW70a]; vertebral centra formed from unfused intercentral half-rings
[AW70a]; parapophyses present [AW70a]; intercentra thickened on each side of
ventral groove to accommodate dorsal aorta [AW70a]; pleurocentra present as
small, rounded elements at ventral end of neural arch [AW70a]; ribs consisting
mostly or entirely of bridge (often incompletely ossified) between parapophysis
and neural arch [AW70a]; anal fin basal with rod-like proximal section [AW70a];
three-lobe caudal fin [CA96] [JA97] [AJ97]; caudal fin , hypochordal lobe
supported by epihemal
spines (articulating with hemal spines), and epichordal lobe by elongated neural
arches [AW70a]; posttemporal present [CA96]; scapulocoracoid with tripodal attachment to
cleithrum [AW70a]; pectoral limb with distal portion of pre-and post-axial elements
flattened [AW70a]; pelvic girdle with long, tapering pubic rami, probably
meeting in cartilaginous symphysis [AW70a]; scales without cosmine [CA96]
[AJ97]; scales round, with median ridge
on inner surface [JA97] [AJ97].
Characters:
some very large (several meters); lateral rostral does not contact prefrontal
[AJ97] [1]; parietals usually long & narrow [AJ97]; postspiracular bones present [JA97] [AJ97];
primitively, lateral extrascapulars are widely separated [JA97]; prefrontal
contacts anterior tectal [JA97]; crista
parotica anteroposteriorly oriented [JA97]; ethmosphenoid cranial
segment elongate [JA97]; vomer posterior processes clasping sides of
parasphenoid [AJ97]; parasymphysial plate small, not contacting coronoid
[JA97]; basal scutes on
unpaired and unpaired fins [AW70a]; vertebral centra formed from unfused intercentral half-rings
[AW70a]; parapophyses present [AW70a]; intercentra thickened on each side of
ventral groove to accommodate dorsal aorta [AW70a]; pleurocentra present as
small, rounded elements at ventral end of neural arch [AW70a]; ribs consisting
mostly or entirely of bridge (often incompletely ossified) between parapophysis
and neural arch [AW70a]; anal fin basal with rod-like proximal section [AW70a];
three-lobe caudal fin [CA96] [JA97] [AJ97]; caudal fin , hypochordal lobe
supported by epihemal
spines (articulating with hemal spines), and epichordal lobe by elongated neural
arches [AW70a]; posttemporal present [CA96]; scapulocoracoid with tripodal attachment to
cleithrum [AW70a]; pectoral limb with distal portion of pre-and post-axial elements
flattened [AW70a]; pelvic girdle with long, tapering pubic rami, probably
meeting in cartilaginous symphysis [AW70a]; scales without cosmine [CA96]
[AJ97]; scales round, with median ridge
on inner surface [JA97] [AJ97].
Image: Tristichopterus at the amazing Paleozoic Fish UK site.
Links: Untitled Document; Eusthenopteron sp.; Eusthenopteron; Hitchcock (French & English); Lobe-Finned Fish; Eusthenopteron and Panderichthyes; Biology 356; Evolution der Fische (German); Palaeontology - Late Devonian fauna from the Hunter Siltstone ....
Mikko's phylogeny shows the phylogeny from [AJ97], which seems to be still more or less curent as of 2005. A PalAss abstract of a 2001 Ahlberg presentation summarizes their position within the Tetrapodomorpha. This work in full is reflected in Ahlberg & Johanson (1998) which is on line at the Australian Museum site. Dennis Murphy has a nice write-up on Hyneria at his Red Hill site.
Notes: [1] a possible synapomorphy, but questioned by [AJ97]. [2] A very nice life image of Hyneria by DW Miller may be found at Famennian.
References: Ahlberg & Johanson (1997) [AJ97], Andrews & Westoll (1970a) [AW70a]; Cloutier & Ahlberg (1996) [CA96], Johanson & Ahlberg (1997) [JA97]. ATW031018.
| Page Back | Unit Home | Page Top | Page Next |
checked ATW051025