Crocodylomorpha
| Palaeos | Crocodilia/ Crocodylomorpha |
|
| Vertebrates | Crocodyliformes |
| Page Back | Unit Home | Unit Dendrogram | Unit References | Taxon Index | Page Next |
| Unit Back | Vertebrates Home | Vertebrate Dendrograms | Vertebrate References | Glossary | Unit Next |
Abbreviated Dendrogram
ARCHOSAURIA | CROCODYLOMORPHA |--Sphenosuchia `--Crocodyliformes |--Protosuchia `--+--Gobiosuchidae `--Mesoeucrocodylia |--Thalattosuchia | |--Teleosauridae | `--Metriorhynchidae `--Metasuchia |--Notosuchidae `--Neosuchia |--Dyrosauridae `--Crocodylia |--Gavialidae `--+--Alligatoridae `--Crocodylidae |
Contents
Overview |
Range: from Late Triassic.
Phylogeny: Crocodylomorpha: Sphenosuchia + *: Protosuchia + (Gobiosuchidae + Mesoeucrocodylia).
Characters: Maxilla shorter than jugal; occipital contacts ventromedial portion of quadrate to enclose carotid and form passages for cranial nerves IX-XI; basisphenoid forms a dorsoventral expanded ("cultriform") process; basisphenoid joint suturally closed; parietal extends dorsally to form ventrolateral edge of trigeminal foramen; antorbital fenestra smaller than orbit; quadrate hollow, with dorsal fenestra; dorsal head of quadrate contacts laterosphenoid; quadratojugal very broad; 2 large palpebrals; skull table flat in temporal region; tail surrounded by osteoderms; ventromedial expansion of coracoid; coracoid anteriorly concave; scapula broadens greatly dorsally (reversed in later forms); pubis at least partly excluded from acetabulum by ant process of ischium.
Links: Crocodylomorpha; NOVA: Outlasting the Dinosaurs; Crocodilian Locomotion (high walk); Clark (1994) cladogram; Crocodylomorpha -- The Dinosauricon.
Discussion: The phylogeny of the Crocodylomorpha is an interesting case study in the status of phylogenetic analysis. Until the mid-1980's the generally accepted phylogeny was essentially Linnaean. Sphenosuchids, Protosuchids, "Mesosuchids," and Eusuchids were believed to form different, and probably successive, suborders of the Order Crocodylomorpha. The exact evolutionary relationships among these groups were studied, but perhaps not a matter of great concern or interest. See, e.g., the treatment by Carroll (1988: 277-83). Then, in that same year, Benton & Clark published their landmark cladistic study which found the Mesosuchia to be paraphyletic. (For reasons to be discussed, this is the scheme that will eventually be adopted here). This was not an unexpected result, and it has widely been accepted as correct. Benton & Clark found that the sphenosuchids and protosuchids were indeed successive branches from the Crocodylomorpha. However, the next major division separated the Thalattosuchia from all other crocs. The Thalattosuchia consist of the well studied marine metriorhynchid and telosaur families (see, e.g. Hua & Buffetaut (1997)). Benton & Clark erected (a) the Mesoeucrocodylia to contain both thalattosuchids and their last common ancestor with living Crocodylia, and (b) the Metasuchia to contain the non-thalattosuchian branch of this clade.
The Thalattosuchia had already been widely recognized as having a close relationships, so this result was reasonably close to expectations. The exact branch point assigned by Benton & Clark, and the separation of, for example, Sebecus from the former "Mesosuchian" mainstream was not as predictable, but again not unreasonable. In fact, it is just this sort of insight that one might expect to get from a computer analysis of the fossil record.
Perhaps unfortunately for croc phylogeny, computers became vastly more powerful over the next few years. Clark (1994) then published a new study using a great deal more morphological data. The resulting cladogram is available, in part, at his home page. A variation is found at the Dinosauricon.
As Clark himself was the first to recognize, something is clearly odd about these results. Clark's analysis reunites all of the Mesozoic marine crocs: not only the Thalattosuchia, but the Dyrosaurs and other groups, which are found to be more basal. Sebecus is now basal to both living crocs and their extinct marine relatives. The problem is that this phylogeny is not only "discordant" with the stratigraphic record, as Clark notes, but it stands the stratigraphic record on its head. The Thalattosuchids are primarily Jurassic and disappear by the early Cretaceous. Dyrosaurs are found from the Upper Cretaceous to the end of the Eocene. Sebecus is restricted to the Eocene of South America. Yet Clark's phylogeny places these groups in the reverse order. Bluntly, it requires more ghost lineages than a Halloween film festival.
What might have gone wrong? Clark himself notes that the results depend on the non-correlation of "longirostrine" characters (the long, thin jaw frequently found in fish-eating forms). As he notes, this assumption is undermined by the presence of the same suite of adaptations in the living gavials, a group which clearly has only distant kinship with Mesozoic marine crocs. Clearly, if most marine piscivores on the croc model are likely to develop the same related group of adaptations, cladistic analysis will tend to group them without regard to actual relationships.
The problem may be more fundamental. Croc phylogeny has a number of features which make it a sort of worst case for cladistic analysis. First, crocs are notoriously conservative. The basic crocodylian design has changed very little in the last 200 million years. As a result, the phylogenetic signal is weak. There is simply very little phylogenetic divergence to detect. Second, the adaptations required by marine life are in fact rather stereotyped. For example, the long, thin piscivorous rostrum, polyphalangy, amphicoelous vertebrae, thickening (pachyostosis) of ribs and vertebrae; development of caudal spines or of a sort of bifurcated caudal "fin", and certain changes in the limb girdles are frequently encountered in all sorts of different organisms making the adaptation to open water life. These changes tend to swamp (perhaps a poor choice of wording in this context) the relatively weak phylogenetic signal in crocs. Third, and to make matters worse, crocs had numerous competitors as large marine predators in the Mesozoic and early Cenozoic: ichthyosaurs, turtles, plesiosaurs and their relatives, mosasaurs, champsosaurs -- not to mention sharks and other predatory fish. This was a contest that the crocs gradually lost. Eventually, crocs were largely restricted to the role of amphibious tropical predators in special environments. As a consequnce, crocs tended to become less marine as time progressed, making the later crocs less specialized than earlier forms. The net result is that Clark's 1994 phylogeny may well be measuring the degree of marine specialization, instead of a real phylogenetic signal, and doing so in circumstances under which the apparent phylogeny reversed the actual pattern of speciation.
If the thesis advanced here is correct, the story of croc phylogeny may have some significant lessons for phylogenetic systematics. Most notably, the assumption that characters are independent cannot be taken for granted. (See glossary entry on parsimony for discussion of this point). The original Benton & Clark (1988) study is preferable because the characters measured are fewer, but more diverse. This increases the chance that the measured attributes are actually independent. The seduction of computing power is in the temptation to measure the same thing many times. The more traits measured, the greater the chance that significant elements are correlated and that the presumption of independence is violated. Phylogenetic systematics has actually shown itself to be remarkably robust, despite the fact that independence cannot be proven in most cases and is probably violated frequently. Nevertheless, independence cannot be assumed away, nor does increasing the size of the character matrix avoid the necessity for validating the assumptions of the study. In fact it may, as here, simply make things worse.
Second, and correlatively, it is neither inappropriate nor unscientific to test the results of cladistics against common sense and stratigraphy. In fact, the assumptions which underlie the cladistic result can usually be tested in no other way. Not only are the assumptions of a study open to question, the quality of the data itself is not always clear. See Ahlberg & Clack (1998) for a recent report in which the inclusion of one poorly understood taxon in the character matrix nearly destroyed the value of an otherwise excellent study. (Indeed, the study is all the more excellent because the authors recognized and dealt with the problem). Unless and until phylogenetic systematics can incorporate the equivalent of a standard error bar in the matrix, it is too easy to claim accuracy when one has only achieved precision.
For reference, the (more or less) complete Benton & Clark (1988) cladogram is as follows [note that this arrangement has now been twice transformed from one format to another and may contain errors] :
Crocodylomorpha |--Pseudohesperosuchus |--Saltoposuchus `--+--Dibothrosuchus |--Sphenosuchus `--+--"Kayenta form" `--Crocodyliformes |--Protosuchidae | |--Hemoprotosuchus | |--Protosuchus+ | `--Kayenta edontosuchus `--+--Odontosuchus `--+--Gobiosuchus `--Mesoeucrocodylia |--Thalattosuchia | |--Pelagosaurus | `--+--Teleosauridae | `--Metriorhynchidae `--Metasuchia |--"Fruita form" `--+--Notosuchus `--+--Baurusuchus |--Libycosuchus `--+--Sebecus `--Neosuchia |--+--Theriosuchus | `--Alligatorium `--+--Dyrosauridae |--Pholidosauridae |--Goniophilidae `--+--Shamosuchus |--Bernissartia `--+--"Glen Rose form" `--Eusuchia |--Hylaeochampsa |--Stromatosuchus |--Leidysuchus `--Crocodylia
Protosuchia: Erythrochampsa (Early Jurassic of South Africa), Nothochampsa (Early Jurassic of South Africa), Orthosuchus (Early Jurassic of South Africa), Protosuchus (Early Jurassic of North America).
Range: upT-upK(?).
Phylogeny: Crocodyliformes: (Gobiosuchidae + Mesoeucrocodylia) + *.
Characters: Reduced upper temporal and antorbital fenestrae; lower canine tooth (like Sphenosuchia) fits into recess between maxilla & premaxilla; basisphenoid larger than basioccipital; extensive ventral contact between quadrate and basisphenoid (quadrate integrated w lateral braincase); pneumatic pterygoid; well-developed secondary palate; quadrate condyles only slightly protruding beyond posterior margin of braincase, and lack of retroarticular process; parietals fused; post-orbital & post-frontals single ossification; post-cranial skeleton like Crocodylia except vertebrae shallowly amphicoelous. Terrestrial forms.
Gobiosuchidae: Gobiosuchus.
Range: upT?
Phylogeny: Crocodyliformes:: Mesoeucrocodylia + *.
Mesoeucrocodylia: Marine & amphibious crocs from the Mesozoic and Cenozoic.
Range: fr lwJ.
Phylogeny: Crocodyliformes:: Protosuchia + *: Thalattosuchia + Metasuchia.
Characters: Posterior extension of palatines (secondary palate); vertebrae amphicoelous; some specialized marine forms with loss of dermal armor, forelimbs as paddles, and thin, elongated snouts.
Links: Paleoecology of the Metriorhynchidae; Crocodilian Biology Database - Locomotion (gallop) (relevance?); DINOSAUR-1995Apr: new Refs; DINOSAUR-1995Oct: Re: Crocodile program on Discovery channel; New ref - Carrier & Farmer; Re: postparietal fenestrae lost; Nathis Krokodillen (Dutch).
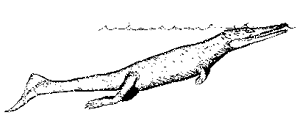 Thalattosuchia:
Pelagosaurus
Thalattosuchia:
Pelagosaurus
Range: lwJ-upEc (lwK??).
Phylogeny: Mesoeucrocodylia: Metasuchia + *: Teleosauridae + Metriorhynchidae.
Characters: highly adapted to aquatic life; long snout; long tail with a tail fin; padle-like limbs; marine.
Links: Paleontology and Geology Glossary: T; paleoecology of the Metriorhynchidae; Untitled Document; Chronologia prochesov (Russian).
Note: Perhaps the only archosaurs fully adapted for marine life. 011113.
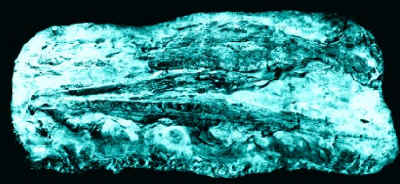 Teleosauridae:
Machimosaurus, Mystriosaurus, Pelagosaurus,
Steneosaurus, Teleosaurus (middle
Cretaceous of North America)
Teleosauridae:
Machimosaurus, Mystriosaurus, Pelagosaurus,
Steneosaurus, Teleosaurus (middle
Cretaceous of North America)
Introduction: One of several families of marine mesosuchian crocodiles.
Range: Early Jurassic to middle Cretaceous of Europe, North America.
Phylogeny: Thalattosuchia: Metriorhynchidae + *.
Characters: The body was long and slim, the back heavily armoured. The jaws extremely narrow and elongated, lined with sharp interlocking teeth from catching fish and belemnites. Forelegs only half the length of the hind limbs, and held flat against the body when swimming. Propulsion came from sinuous movements of the body and tail. Not highly specialized for marine life and may have retained amphibious habits.
Links: PANGEA, Untitled Document; Information: Krokodil (German); Teleosauridae; juras2.html (Spanish); Bone histology as a clue in the interpretation of functional ... (JVP abstract); SUI RESTI DI COCCODRILLO DEL ROSSO AMMONITICO VERONESE DI SASSO ... (article in Italian); reptiles (table); PANGEA (table); Fauna der Oberen Jura Deutschlands; Information- Krokodil; ATW031027.
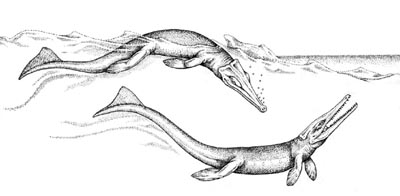 Metriorhynchidae: Geosaurus,
Metriorhynchus, Teleidosaurus.
Metriorhynchidae: Geosaurus,
Metriorhynchus, Teleidosaurus.
Range: Early to Middle Jurassic of Europe and South America.
Phylogeny: Thalattosuchia: Teleosauridae + *.
Characters: Skull dorsoventrally flattened (quick lateral attacks); skull highly pneumatized; nares are not raised; osteoscutes absent; forelimbs with hydrofoil shape; generalist swimmers; epipelagic stalking predators
Links: paleoecology of the Metriorhynchidae; Untitled Document; cocodrilos marinos (Spanish. Best on the Web); PANGEA (Italian); Pelagosaurus typus (French); Information: Krokodil (German). 020317.
| Page Back | Unit Home | Page Top | Page Next |
checked ATW031027
{kind=link}