| Palaeos | Coelurosauria | |
| Vertebrates | Alvarezsauridae |
| Page Back | Unit Home | Unit Dendrogram | Unit References | Taxon Index | Page Next |
| Unit Back | Vertebrates Home | Vertebrate Dendrograms | Vertebrate References | Glossary | Unit Next |
|
Abbreviated Dendrogram
DINOSAURIA |--ORNITHISCHIA `--+--SAUROPODOMORPHA `--THEROPODA | `--COELUROSAURIA |--Tyrannosauroidea `--+--Ornithomimosauria `--+--Maniraptora |--Therizinosauroidea `--Metornithes |--Alvarezsauridae | |--Alvarezsaurus | `--+--Patagonykus | `--Mononykinae |--Oviraptorosauria `--Paraves |--+--Troodontidae | `--Dromaeosauridae `--AVES |
Contents
Index |
Editor's note: this and the following page were originally included under the unit Aves. They have been relocated to the present unit following current understanding, which places the Alvarezsauridae stemward of (i.e. more basal than) true birds. However the text has not yet been revised. MAK120309
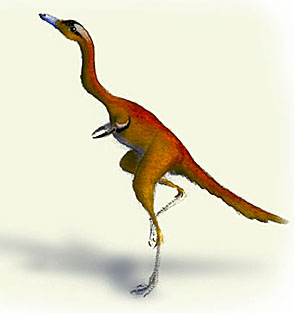
Some of the strangest of early birds are the alvarezsaurs. These creatures were plainly flightless, and the nature of their flightlessness requires some special comment. The Alvarezsauridae are, conceptually, the very opposite of Archaeopteryx. The weirdness of Archaeopteryx is that it was plainly designed to fly, but had almost no bird-like features: no keeled sternum, little or no fusion of wrist or ankle bones; not much in the way of special skull adaptations, and so on. By contrast, the Alvarezsaurids have all these and more. The weirdness of the Alvarezsaurids is that, whatever they were adapted for, it was plainly not flight. One can envision any number of strange and wonderful keratinous or cartilaginous appurtenances for those short, but heavily muscled, single-fingered arms. However, it is beyond conception that any of them could ever have permitted Mononykus to fly.
 As
Henry Gee points out (in his usual pugnacious, but always eloquent, style)
flight is very much a mixed blessing. It confers enormous advantages. However,
at the same time, it demands an enormous dedication of energy and an enormous
commitment of anatomical resources that might find "better" uses.
Giving up on flight confers a large energy bonus and so, "the world is
full of flightless birds." Gee (1999: 199);
see also Feduccia (1999: 231).
As
Henry Gee points out (in his usual pugnacious, but always eloquent, style)
flight is very much a mixed blessing. It confers enormous advantages. However,
at the same time, it demands an enormous dedication of energy and an enormous
commitment of anatomical resources that might find "better" uses.
Giving up on flight confers a large energy bonus and so, "the world is
full of flightless birds." Gee (1999: 199);
see also Feduccia (1999: 231).
But this may not be quite the same thing. The world is full of flightless birds which share certain characteristics. Feduccia (1999: 289). Cenozoic flightless birds do not have a keeled sternum. In fact, this is virtually the first anatomical "event" that occurs when flight is lost. Other relatively predictable adaptations are the development of an obtuse angle between the scapula and coracoid and the loss of the furcula. Since flightless birds are not constrained to minimize weight to the same extent as flying forms, they typically become, not only larger, but fatter at any given size. Feduccia (1999: 258-68). Modern flightless birds (mostly, but not entirely, ratites) also typically evolve in isolated locations, often islands where competition for a variety of niches is low. They have not competed well with other forms and tend to disappear or become marginalized fairly quickly once competition is introduced. Like adventitious plants, birds can fill ecological vacuums very quickly but are not necessarily robust competitors over the longer term.
Alvarezsaurids, by contrast, seem to represent a group that enjoyed considerable, long-term success in both Mongolia and Argentina, regions which are located on large continental land masses and were almost as remote from each other in the Cretaceous as they are today. Certainly alvarezsaurids did not have to compete with advanced rodents (the bane of many more recent flightless bird species), but their very broad geographic dispersion is notable. They plainly retained a keeled sternum and a robust pectoral girdle. The scapula - coracoid angle is problematic, but appears to have been only slightly obtuse in Mononykus, the most derived member of the clade. The clavicles (so far as I have been able to determine) are not known and the furcula may have disappeared. However, the alvarezsaurids remained, insofar as one can judge from bones, notably scrawny over the considerable size range from Parvicursor to Mononykus and did not put on weight like an elephant bird (Aepyornithiformes), or even a kiwi (Apterygiformes).
On the whole, the alvarezsaurids may point to a somewhat different pattern of evolution. The most striking difference is that the arms, while not useful for flight, were plainly useful for something. When neornithines become flightless, the pectoral apparatus is very quickly reduced -- a matter of generations, not megayears. Feduccia (1999: 266). The selective advantage in "bulking down" is strong because the flight muscles comprise an average of 20% of the body mass of even highly modified, flight-adapted neornithine birds. Even without the added weight of the pectoral girdle supporting flight, this represents an enormous metabolic commitment, and the energetic reward for losing it is a powerful selective advantage. Despite this selective pressure, alvarezsaurids seem to have lost the ability to fly while retaining much of the anatomical baggage that made flight so expensive. One of the principal advantages of flightlessness is the chance to get fat and lazy (i.e., to reduce basal metabolism and accumulate fat reserves). From all appearances alvarezsaurids did neither.
At this point, the perceptive essayist would introduce some startling insight into the lives of alvarezsaurids which would explain their strangeness. Unfortunately, nothing of the sort is on offer here. However, there is one possible evolutionary parameter that might be considered, given the exceedingly odd pattern of alvarezsaurid evolution. Perhaps the alvarezsaurids did not take full advantage of the metabolic advantage of flightlessness because there was less advantage to be had. If dinosaurs did not develop full endothermy until after the development of more advanced avian adaptations, then the energetics of secondary flightlessness simply didn't require the immediate loss of the flight apparatus and reduction of basal metabolism. Flight would be a short-term, anaerobic, hyper-exertion. The loss of flight would free up anatomical resources and open up "design" possibilities. It would not, however, involve as much in the way of immediate metabolic savings. The reduction in selective pressure could, in turn, slow down the process of de-adaptation for flight long enough to permit selection for some other forelimb function.
By contrast, it is noteworthy how few flightless neornithines have made any significant use of their forelimbs. Perhaps the only exceptions are auks, penguins and other birds which "fly" under water. The exceptions support the rule. It is perfectly possible to use wings for both flying and swimming (some petrels, razor-bills). Thus diving wings are an unremarkable adaptation because the development of such wings does not involve the same kind of geologically instantaneous vestigialization of the flight apparatus. The latter creates few opportunities for retooling.
Such a paradigm would go some way toward explaining one of the open questions about flight in Archaeopteryx. Flight, in efficient, neornithine birds, is terribly expensive. It is so expensive that the ability to fly is subject to enormous negative selection whenever circumstances permit survival without it. In the competition between the flying and non-flying descendants of island colonizers, the non-flyers seem to triumph in the blink of a geological eye. But, if this be true of efficient, highly derived neornithine flyers, how could a relatively inefficient, proto-archaeornithine almost-flyer ever overcome this negative selection and develop flight in the first place?
The answer that comes most easily to hand is that the selective advantage of flightlessness may have been less clear. The barrier might be lower because the energetic cost of flight did not include maintaining huge flight muscles at a constant internal temperature, with all that this entails. The proto-flyer's basal metabolism was already low. The gain from giving up flight muscle was not as compelling a selective argument as it later became. Indeed -- to crawl even further out on this limb -- it might be more sensible to suppose that the size and strength of the flight muscle later drove full Neornithine homeothermy, rather than the reverse. A bird fully committed to flight gains advantages by endothermy which would more than fully compensate for the energetic cost.
That half of the equation may be left for another day and another taxon. For the alvarezsaurids, who manifestly did lose flight, the net result of primitively lower metabolism would be what is actually observed: the flight apparatus was not lost, because it had not been all that expensive in the first place. Selection against flight was not the break-neck affair it is believed to be in the case of isolated island neornithines. Rather, the conversion was a slow and statistical process in which the flight apparatus was gradually converted to another use.
Now if we could only determine what that use was ...
Mononykus olecrans image courtesy of Daniel Bensen and T. Michael Keesey, whose scientific and artistic work, including this image or versions of it, may be accessed, respectively, at OPUS: Dinosaur and The Dinosauricon. (© as stated at those sites).
Metornithes: definition: Alvarezsaurus + Neornithes. Padian et al. (1999) [C95]
Range: from the Early Cretaceous.
Phylogeny: Aves: Archaeornithes + *: Alvarezsauridae + (Confuciusornithidae + Ornithothoraces)
Characters: $ loss of "jugular" (jugal?) postorbital process [F+98]; $ medial otic process of quadrate articulates with the prootic [F+98]; quadrate articulates with prootic and squamosal; $ prominent ventral processes on cervicodorsal vertebrae [C95]; vertebral neural canal dorsoventrally wide relative to centrum depth [P+93]; $ large rectangular sternum [P+93] [C95]; $ sternum with ossified carina [C95] [P+93]; $ ventral tubercle of humerus projects caudally and is separated from the humeral head by distinct "capital incision" (neck?) [F+98]; ulnar distal condyle present; $ fused carpometacarpus [F+98] [P+93, but ambiguous] [C95]; $ prominent antitrochanter on ilium (but wouldn't this be primitive?) [F+98] [P+93]; ischium > 2/3 length of pubis; $ obturator process on ischium rudimentary or absent [F+98]; $ pubic apron transversely narrowed with pubic symphysis restricted to distal one-third of shaft [F+98]; compare no pubic symphysis [P+93]; $ loss of pubic foot [F+98] [P+93]; femur with undivided trochanteric crest [P+93]; femur with distal condyles nearly confluent below popliteal fossa [P+93]; $ fibula does not reach tarsus [P+93].
Links: So here it is... my paper, open for discussion (very long; try to view it in maximum window breadth) (strange, but possible); Re: I'm back...; Re: Coelurus a maniraptoran (for how long?); THEM EARLY BIRDS; Re: Carpometacarpus.
References: Chiappe (1995) [C95]; Forster et al. (1998) [F+98]; Perle et al. (1993) [P+93].
Note: [1] It is interesting that the basic structure of the wing itself came first, followed by the reorganization of the limb girdle and musculature necessary to make really effective use of the wing in flight. [2] The tendency now is to put alvarezsaurids somewhere back in basal Maniraptora. See, e.g., Coelurosauria -- The Dinosauricon. In that case, its hard to tell what this clade contains. However, Mikko's site has a different definition (Neornithes > Archaeopteryx), attributed to Chiappe, which I like much better and which would preserve the utility of this clade. ATW020809.
Update: since the above was written, the evolutionary relationships of this clade has been totally revised, with all recent studies unanimously placing the Alvasauridae stemward of Archaeopteryx and Aves (Zhang et al 2008, Zanno et al 2009, Xu et al 2011, Lee & Worthy 2011). The following 2005 comment by Tim Williams, from the Dinosaur Mailing List pretty much sums up the resulting phylogenetic inversion:
"...The discovery and or inclusion of one new specimen or one new taxon can shift taxa around and play havoc with topology. I seem to remember that _Mononykus_ once looked quite secure as a basal avialan more derived than _Archaeopteryx_, and there were plenty of papers arguing this point. Perle et al. (1993) erected the clade Metornithes under the assumption that _Mononykus_ (=_Mononychus_) was a bird more derived than _Archaeopteryx_. Subsequently, _Mononykus_ (and other alvarezsaurs) fell out of the Avialae. Now, with its current definition, Metornithes is bigger (more inclusive) than Avialae! If Metornithes had been defined as "all descendents of the most recent common ancestor of _Mononykus_ and _Passer_, but not _Archaeopteryx_", then Metornithes would disappear in the event that _Mononykus_ was found to be more basal than _Archaeopteryx_. In other words, once the avialan hypothesis for _Mononykus_ was no longer supported, the clade Metornithes (which was erected in the context of _Mononykus_ being a bird) would also expire."
For Paul 1988 and 2002, all maniraptors are neoflightless descendents of archaeopterygians. This is in agreement with Perle et al. 1993, but not with either earlier or more recent (e.g.Senter 2007) cladistic analyses of theropods and coelurosauria. In the standard hypothesis (birds evolved from troodont or dromaeasaur-like maniraptors), the original definition of Metornithes by Perle et al. (1993) as the clade containing Mononykus and Ornithothoraces, but not Archaeopteryx and non-avian theropods, becomes invalid. Because Mononykus and Alvarezsaurus belong to the same clade, Chiappe's (1995) definition of Metornithes as Mononykus + Neornithes is and Padian et al. (1999) definition are synonymous. Mickey Mortimer makes alvarezsaurids the sister taxon to the ornithominosaurs, and hence Metornithes synonymous with Maniraptoriformes. This differs from the consensus of current papers which place the Alvarezsaurs fully in the maniraprora, although it does recall Greg Paul's ornithomimids as neo-flightless birds. In any case, the close similarity between troodonts, dromaeasaurs, oviraptors, archaeopterygids, and alverezsaurs, with alverezsaurs in some regards appearing more bird-like than Archaeopteryx, shows how similar all these proto-birds were, as well as the nature of mosaic evolution and the way in which convergence, reversals, and other evolutionary artifacts make it difficult if not impossible to determine the extact relationships among the various proto-birds and dino-birds. MAK120306.
Alvarezsauridae: 1-2 m flightless birds with small arms. Elaphrosaurus gautieri? (see Mortimer), Rapator? (see Rapator ornitholestoides, large Aussie dinosaur -- or bird?)
Range: Late Cretaceous of South America, Mongolia & possibly Africa.
Phylogeny: Metornithes: (Confuciusornithidae + Ornithothoraces) + *: Alvarezsaurus + (Patagonykus + Mononykinae).
Characters: prenarial snout absent; upper jaw moveable on skull (i.e., without opening mouth); prefrontal large with broad orbital flanges; no postorbital-jugal contact; movable joint between quadratojugal and quadrate, no quadratojugal-squamosal contact; separate squamosal and braincase articulations of quadrate, tympanic recess opens within columellar recess (not paroccipital process); triradiate palatine; teeth present on dentary & premaxilla; maxilla without teeth; teeth with unserrated crowns; shortened vertebral spines; dorsal centra with very deep neural canal, approaching 50% centrum height; $ sacrals procoelous [N97]; $ posterior sacral centra ventrally keeled and laterally compressed [N97]; synsacrum with ~7 vertebrae; caudal count < 25-26; 1st (and distal?) caudal centrum (centra?) with ventral keel; caudals procoelous; tail moderately long (?); proximal chevrons long; caudal zygapophyses reduced & short; caudal hemapophyses not reduced; sternum rectangular; carinate sternum (carina small); glenoid does not face laterally; humerus very short; fused carpometacarpus; arms short, probably heavily muscled; humerus stout; in M., large olecranon process and single large manual digit; $ robust ungual on manus I with quadrangular articular surface [N97]; manual unguals with flattened ventral surfaces and distally displaced flexor tubercles; supraacetabular crest ends above acetabulum; pelvis with prominent antitrochanter; $ iliac pubic peduncle slender [N97]; ilium pubic peduncle projects anteroventrally; iliac blades meet in the midline; ischium > two-thirds length of pubis; no contact between distal ends of ischia; legs very long, esp. in M.; fibula splint-like and incomplete; no medial fossa on proximal end fibula; primitively, at least one digit reversed (?).
Links: DinoData: Mononykus; DinoData: Shuvuuia; Link (Japanese); Mononykus (Nakasato); Nakasato ...Mononykus (forelimb); National Geographic ...1996; Lectures 19-20; Bonezone - Mark Norell; Alvarezsauria -- The Dinosauricon; polyphyletic Alvarezsauria assemblage; Arctometatarsalia and Alvarezsauria; \Elaphrosaurus\ gautieri identified as an ...... (and follow-ups thereto).
References: Novas (1997) [N97]. ATW011029.
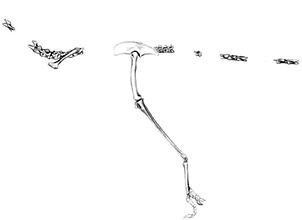 Alvarezsaurus:
Bonaparte 1991. A. calvoi Bonaparte 1991.
Alvarezsaurus:
Bonaparte 1991. A. calvoi Bonaparte 1991.
Range: Late Cretaceous (Coniacian to Santonian) of South America.
Phylogeny: Alvarezsauridae: (Patagonykus + Mononykinae) + *.
Characters: ~2m & perhaps 20 kg; cervical pleurocoels present; last sacral & caudal vertebrae procoelous [N97]; caudal neural spines very low; tail very long; distal caudal elongate [N97]; single digit on forelimb (??); supraacetabular crest absent [N97]; femur 4th trochanter absent [N97]; fibula articulates (fuses?) distally with tibia [N97]; pes not arctometatarsalian [N97].
Notes: as [N97] points out, the forelimb of Alvarezsaurus is actually unknown, so that the peculiar alvarezsaurid claws may or may not be a synapomorphy of the Alvarezsauridae.
Links: Alvarezsaurus -- The Dinosauricon; Chamworks Dinosaurs & Such: Alvarezsaurus calvoi; DinoData Dinosaurs A090 ALVAREZSAURUS calvoi; ALVAREZSAURUS; Alvarezsaurus (Dutch); Alvarezsaurus.htm (Spanish); Alvarezsaurus page in The Natural History Museum's Dino Directory; DinoDictionary.com | A - Dinosaurs Page 2.
References: Novas (1997) [N97]. ATW020810.
 Patagonykus:
P. puertai Novas 1996.
Patagonykus:
P. puertai Novas 1996.
Range: Late Cretaceous (Turonian) of South America
Phylogeny: Alvarezsauridae:: Mononykinae + *.
Characters: cervical vertebrae with pleurocoels present; $ dorsal
vertebrae postzygapophyses with curved, tongue-shaped lateral margin [N97];
dorsal vertebrae without pleurocoels [N97]; dorsal vertebrae with accessory
hyposphene-hypantra articulations; hyposphenes present as deep and wide ridge
between postzygapophyses [N97]; neural spines dorsally flat and elongated
anteroposteriorly [N97]; dorsal parapophyses & diapophyses directed (i.e.
the ribs were directed) laterally & slightly dorsal [N97]; $ dorsal, sacral
& caudal vertebrae with bulge at caudal base of neural arch [N97]; dorsal, sacral
& caudal vertebrae procoelous [N97]; probably 5 sacrals [N97]; sacrals
hollow, with thin periosteum [N97]; sacral centra narrow & deep [N97];
sacral transverse processes directed laterodorsally [N97]; posterior sacrals
with ventral keel [N97]; anterior caudals similar to sacrals [N97]; caudal
vertebrae with postzygapophyses much longer than in dorsals; distal caudals
dorsoventrally flattened, with distinct grooves on ventral surface [N97]; $
coracoid with transversely narrow humeral facet [N97]; coracoid posterior
process long, flat & ventrally 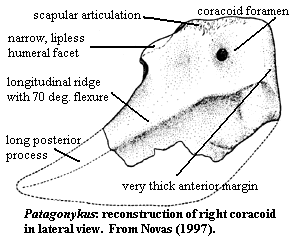 deep
[N97];anterior margin is very thick [N97]; coracoid with long ridge, marking
flexure of ~70 º [N97]; forelimb very similar to Mononykus [N97];
humeral head convex, long & narrow (wide range of motion?) [N97]; humeral
head may have been continuous with deltopectoral crest [N97]; $ humerus internal
tuberosity cylindrical & broadening distally [N97]; ends of both head and
tuber smooth, convex & ringed with attachment scars [N97]; $ humerus
entepicondyle conical & projects strongly medially [N97]; humeral
ectepicondyle separated from condyle by marked notch and strongly projecting
almost distally [N97]; humerus with single, large, spherical distal condyle
[N97]; ulna as in Mononykus, but even more robust [N97]; carpometacarpus
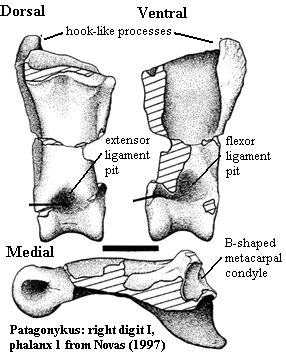
has semilunate carpal fused to large MtI [N97]; $ manual digit I phalanx 1 with
proximomedial, ventrally-directed hook-like process(es) [N97]; digit is
dorsoventrally flattened; supraacetabular crest prominent & projects
laterally, but is restricted to dorsal part of acetabulum [N97]; ilium ischiac
pedicel with raised antitrochanter [N97]; pubis with small obturator
notch distal to articulation for ischium (with corresponding notch on ischium)
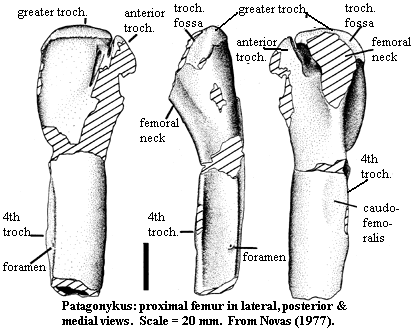
[N97]; pubic symphysis and boot present [N97]; femoral anterior (= lesser)
trochanter projects up to level of greater trochanter, but is separated by deep
cleft [N97]; cleft continues as excavation separating anterior trochanter from
femoral head [N97]; 4th trochanter present, with prominent foramen and slight
depression for insertion of m. caudofemoralis longus [N97]; proximal
femoral shaft laterally flattened [N97]; $ "ectocondylar tuber of femur
rectangular shaped in distal view" [N97]; tibia proximal articular surface
almost flat [N97]; tibia with strong, but short cnemial crest [N97]; tibia with
outer and inner condyles on posterior side, separated by deep, narrow notch
[N97]; distal tibia closely appressed to astragalus but not fused [N97]; fibula articulates distally with
deep
[N97];anterior margin is very thick [N97]; coracoid with long ridge, marking
flexure of ~70 º [N97]; forelimb very similar to Mononykus [N97];
humeral head convex, long & narrow (wide range of motion?) [N97]; humeral
head may have been continuous with deltopectoral crest [N97]; $ humerus internal
tuberosity cylindrical & broadening distally [N97]; ends of both head and
tuber smooth, convex & ringed with attachment scars [N97]; $ humerus
entepicondyle conical & projects strongly medially [N97]; humeral
ectepicondyle separated from condyle by marked notch and strongly projecting
almost distally [N97]; humerus with single, large, spherical distal condyle
[N97]; ulna as in Mononykus, but even more robust [N97]; carpometacarpus
has semilunate carpal fused to large MtI [N97]; $ manual digit I phalanx 1 with
proximomedial, ventrally-directed hook-like process(es) [N97]; digit is
dorsoventrally flattened; supraacetabular crest prominent & projects
laterally, but is restricted to dorsal part of acetabulum [N97]; ilium ischiac
pedicel with raised antitrochanter [N97]; pubis with small obturator
notch distal to articulation for ischium (with corresponding notch on ischium)
[N97]; pubic symphysis and boot present [N97]; femoral anterior (= lesser)
trochanter projects up to level of greater trochanter, but is separated by deep
cleft [N97]; cleft continues as excavation separating anterior trochanter from
femoral head [N97]; 4th trochanter present, with prominent foramen and slight
depression for insertion of m. caudofemoralis longus [N97]; proximal
femoral shaft laterally flattened [N97]; $ "ectocondylar tuber of femur
rectangular shaped in distal view" [N97]; tibia proximal articular surface
almost flat [N97]; tibia with strong, but short cnemial crest [N97]; tibia with
outer and inner condyles on posterior side, separated by deep, narrow notch
[N97]; distal tibia closely appressed to astragalus but not fused [N97]; fibula articulates distally with
 tibia [N97]; fibula not in contact
with calcaneum (contra? [N97]); astragalus fused to calcaneum [N97];
astragalus ascending process wide, but laminar & very (<1 mm) thin [N97];
metatarsals II & III partially fused [N97]; pes III not arctometatarsalian (vs. Mononykus)
[N97].
tibia [N97]; fibula not in contact
with calcaneum (contra? [N97]); astragalus fused to calcaneum [N97];
astragalus ascending process wide, but laminar & very (<1 mm) thin [N97];
metatarsals II & III partially fused [N97]; pes III not arctometatarsalian (vs. Mononykus)
[N97].
Note: [1] very fragmentary remains of a second, unnamed alvarezsaur were found with Patagonykus. [N97]. [2] It is very, very difficult to make sense of this arm -- even more difficult than in Mononykus. The humeral head seems to articulate comfortably with the coracoid facet only if the arm were sticking straight out sideways. In this position, it is hard to see how the humerus could be rotated anteriorly, because of the long contact between coracoid and humeral head. If this is correct, the net result is that Patagonykus could only flap its arms up and down. The short, but powerful, claw and forearm would then appear to move -- rather uselessly -- in a sort of radial plane, between pointing anteriorly to pointing straight out sideways. Perhaps the creature makes better sense if we reconstruct it as a much fatter beast. This is also suggested by the dorsolateral angle of the rib articulations. The coracoids can then be placed on a broad, muscular breast and face more anteriorly than laterally. If we then rotate the right coracoid counterclockwise, the up-and-down motion becomes an in-and-out motion and the scapular blade points dorsolaterally, which looks about right and makes much better mechanical sense.
Links: DinoData Dinosaurs P049 PATAGONYKUS puertai; \Elaphrosaurus\ gautieri identified as an ......; PATAGONYKUS; Extinct Birds; The Dinosauria by DinosØMP; Patagonykus -- The Dinosauricon; The Theropod Ancestry of Birds; The Journal of Vertebrate Paleontology; Bruno Hernandez's Paleo Gallery @ Prehistorics Illustrated page 08; ALVAREZSAURIDAE (German); El mundo perdido de la Patagonia (Spanish); Argentina On View (Spanish); MEA592D Dinosaur Osteology: Lecture 6.
 References: Forster et al. (1998),
Novas (1997) [N97].
ATW020810.
References: Forster et al. (1998),
Novas (1997) [N97].
ATW020810.
| Page Back | Unit Home | Page Top | Page Next |
checked ATW050717