| Palaeos: Mesozoic | Late Triassic epoch | |
| Triassic Period | Carnian Age - 2 |
| Page Back | Back: Ladinian | Back: Middle Triassic | Up: Carnian 1 | Unit Home |
| Page Next | Next: Norian | Next: Early Jurassic | Timescale |
| Mesozoic Era
Triassic Period Early Triassic Epoch Middle Triassic Epoch Anisian Age Ladinian Age Late Triassic Epoch Carnian Age Norian Age Rhaetian Age Jurassic Period Cretaceous Period |
Introduction
The Geography of the Carnian The Climate of the Carnian Stratigraphy Plants Invertebrates Tetrapods - Land Faunal Stages A Carnian Bestiary Links |
Pelorocephalus | Metoposaurus | Leptopleuron | Shonisaurus | Henodus | Brachyrhinodon | Malerisaurus | Trilophosaurus | Hyperodapedon | Paleorhinus/Parasuchus | Angistorhinus | Saurosuchus | Ornithosuchus | Stagonolepis | Postosuchus | Erpetosuchus | Scleromochlus | Pisanosaurus | Herrerasaurus | Saltopus | Placerias and Ischigualastia | Exaeretodon | Microconodon and Dromatherium | Gondwanadon and Adelobasileus
 Dinosaurios y animales prehistoricos argentinos/Ischigualasto fauna |
|
Pelorocephalus ischigualastensis Bonaparte, 1975 was a chigutisaur, a type of temnospondyl amphibian known only from Gondwana/South Pangea. The late Carnian was the high point of these creatures; as well as several species from Argentina, there
are two genera from the Upper Maleri Formation of India. These specialized short-headed amphibians were one of the very few labyrinthodonts to survive the end Triassic extinctions. During the Jurassic and Middle Cretaceous crocodiles are unknown and giant Chigutisaurids (3 to 5 meters) flourished in south-east Gondwana.
Pelorocephalus was originally known as Chigutisaurus, the genus from which the family derives its name. But whereas Chigutisaurus was named in 1948, the Pelorocephalus
moniker goes back a further four years, to 1944, and hence has priority, under the formal rules of Linnean (binomial) nomenclature
Carnian species:
|
 |
|
The Carnian temnospondyls were every bit as diverse as their Ladinian predecessors. In addition to the familiar mastodonsaurids (Cyclotosaurs) there were also several newcomers - the big metoposaurs and small latiscopids (both included under the same superfamily). No plagiosaurs are known from the Carnian but they must surely have been present, since their remains are found in both Ladinian and Norian deposits. The metoposaurs are one of the Carnian success stories. They appear to come seemingly from nowhere (although possible early forms are reported from the Induan [J.S. Steyer 2002) and attain near-global distribution. They were large aquatic forms (2 to 2.5 meters in length) with huge flattened skulls, very much like the Capitosauridae/Mastodonsauridae, but differing in details and proportion of the skull. The European (North Central Pangea) metoposaurs differ in small details from the North American and Indian (north-western and central southern Pangean) forms, which differ again from the central Pangean (Morocco) species. Although these have all been given different generic names, they are very similar and probably could equally be retained under Metoposaurus. Shown here is Buettneria perfecta, a metoposaur whose fossil remains are known from both the Chinle Group and Newark Supergroup, North America (then west and central equatorial Pangea). Length of skull about 40 cm; overall length about 2 meters. Like almost all metoposaurs (apart from the aberrant small terrestrial Apachesaurus) these animals probably spent much of their time submerged in ponds, rivers, and swamps. |
from Kuhn 1969 after Huene 1912 (skeleton) and 1956 (skull) |
||
|
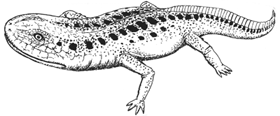
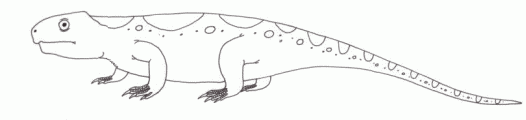
Leptopleuron lacertinum, known from the Lossiemouth Beds, Elgin, Scotland, represents yet another representative of the primitive (parareptile) reptilian herbivores called procolophonids. These stocky herbivorous desert "lizards" were a ubiquitous component of many Triassic terrestrial ecosystems, especially the more arid environments; which during late Triassic Pangea meant the higher and more desert-like latitudes [Olsen et al]. Leptopleuron was a fairly advanced form, as indicated by its broad spiny skull, and would seem to be closely related to the late Norian to Rhaetian Hypsognathus, as well as to several other little known Norian forms [Sues et al 2000]. This could all be placed in the family Leptopleuroninae Ivakhnenko, 1979. It would seem then that during the late Triassic the leptopleuronines completely replaced the older and less specialized procolophonines |
 drawing © 2001 Vince R Ward - Prehistoric Pages |
|
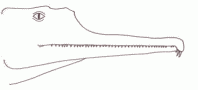
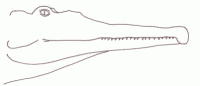
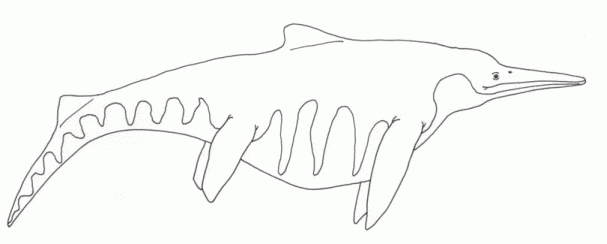
During the Carnian epoch the Ichthyosaurs continued to flourish, a number of forms existing alongside each other. These have traditionally been classified under the families Shastasauridae and Shonisauridae, although more recent research has the shown these to be paraphyletic or even polyphyletic groupings, part of the clade Shastasauria, and the relationships between these various types not well known, owing to fragmentary material in the case of many forms. These were large animals, and grew progressively more so; "Shastasaurus" beginning at 3 meters and increasing to 10 meters in stratigraphically younger forms [Zittel, 1932 p.280]. Shonisaurus was even larger but
exhibited a similar trend [Camp, 1980], so that the latest Carnian forms reached more than 15 meters. While one related
Norian form was about 23 meters along, the largest Mesozoic marine animal known. It is possible that this rapid increase in size was encouraged by the vacant ecological niche left by the extinction of earlier large marine reptiles Cymbiospondylus and Paranothosaurus, but the
shastasaurs grew much larger than their predecessors.
|
 illustration by Steve Kirk, © from Encyclopaedia of Dinosaurs and Prehistoric Animals, Barry Cox, R.J.G.Savage, Brian Gardiner, Dougal Dixon, |
|
Henodus chelyops Huene 1936 is known from a number of skeletons from the Gipskeuper (Early Carnian) of Tübingen, southern Germany. The only known non-marine placodont, it inhabited brackish or possibly freshwater lagoons, and may have fed on fishes and crustaceans as well as molluscs. As with the Ladinian Placochelys, it was a member of the
Placodonta that had evolved to look like turtles, with a broad armoured carapace. The back and belly were covered with an armour of irregular, many-sided bony plates, the whole being completely covered with horn. The square toothless snout was equipped in life with a strong horny beak, another parallel with a modern turtle. Originally it was considered to be an advanced species, but recent research indicates that Henodus may actually represent a primitive form, and its late appearance in the fossil record explained by the fact that it was restricted to a peculiar environment that is not preserved in older rocks of the same region. [Rieppel and Zanon 1997). Henodus reached an overall length of about 1.2 meters.
|
 drawing © 2001 Vince R Ward - Prehistoric Pages slightly modified |
|
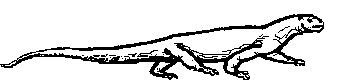
Brachyrhinodon taylori is one of the oldest known sphenodonts, a group that filled the important lizard ecomorph in the late Triassic and throughout the Jurassic, when true lizards remained less common. This little animal is remarkable for the brevity of the snout, which would seem to represent a specialized adaptation. It is known from poorly preserved casts in the Elgin sandstone, which makes exact relationships difficult to determine, and has been placed in its own subfamily Brachyrhinodontinae [Kuhn 1969], related to the Polysphenodontinae [Fraser 1986], or placed with Polysphenodon in a clade that includes Clevosaurus [Wu, 1994]. In any case, these little reptiles had already become a part of the small insectivore guild, and were clearly more widespread and diverse then their fragmentary remains indicate |
 llustration by Gregory S Paul |
|
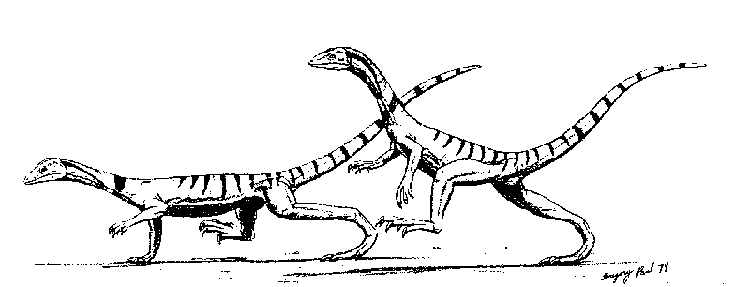
Malerisaurus was a lizard-like prolacertiform, superficially not unlike the Induan Prolacerta, which averaged about 1.2 meters in length. It probably had rather similar habits to a large tropical lizard. The hindlimbs were larger than the front legs, indicating that, like Permo-Triassic Younginoformes and the modern fill-necked lizard of Australia, these animals could rear up on their hind limbs for an extra burst of speed. Feeding on invertebrates and small vertebrates, Malerisaurus itself fell prey to larger hunters. Two specimens of M. robinsonae were found preserved in the stomach cavity of a Parasuchus phytosaur. These animals were widely distributed during late Triassic times. Malerisaurus robinsonae Chatterjee 1980, illustrated here, is known from the early Late Carnian [Otischalkian] Maleri Formation, of the Pranhita-Godavari Valley, India. A very similar species, M. langstoni Chatterjee, has been found in strata of the contemporary Dockum Formation of Texas |
 llustration (such as it is) by M. Alan Kazlev |
|
Trilophosaurus buettneri were large (length about 2.5 meters) herbivorous reptiles, with a body in the form of a large lizard, but with an incongruous-looking small turtle-like heads and a toothless beak. These sprawling reptiles appeared suddenly in the late Carnian and flourished briefly before dying out, although some small Norian forms from Europe may be related. Trilophosaurus is from the Otis Chalk, Dockum formation, Chinle Group, of Texas, USA (equatorial west Pangea) early Late Carnian [Otischalkian age]. Trilophosaurus this was at one time considered a euryapsid, owing to similarities of the skull to the aquatic reptiles like ichthyosaurs and plesiosaurs. In more recent decades, it has been conclusively shown that they are actually archosauromorphs, and their nearest relatives are the lizard-like Prolacertiformes (e.g. Malerisaurus) and stock-bodied rhynchosaurs (e.g. Hyperodapedon) |
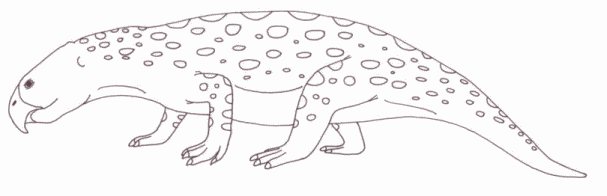 drawing © 2001 Vince R Ward - Prehistoric Pages |
|
The Rhynchosauridae were a family of purely Triassic archosauromorphs that became
particularly important during the Carnian age. All Carnian forms belong to the advanced subfamily Hyperodapedontinae, and most can be included in the genus Hyperodepedon Huxley, 1859. These were medium-sized terrestrial herbivores (Skull length about 20 cm; average overall length 1.3 to 1.5 meters), with stocky barrel-like bodies, broad heads and beaked jaws. There were two broad tooth plates on each side of the upper jaw, and these had a groove that matched the single tooth row on each side of the lower jaw, giving a very efficient cutting action. The heads were also much broader than their predecessors, to enable larger rows, and have the most advanced dentition of any rhynchosaurs. |
drawing © 2001 Vince R Ward - Prehistoric Pages; skull from A. H. Müller, Lehrbuch der Paläozoologie |
||
|
The fish, amphibians and smaller reptiles of the Carnian rivers and swamps frequently fell prey to animals that bore a remarkable resemblance to modern crocodilians. These were the rather inappropriately-named phytosaurs ("plant saurians" because the original describer believed that the mud fillings between the teeth sockets were actually thick
herbivore teeth). These were large armoured semi-aquatic archosaurs with a primitive ankle structure and sprawling posture (either these were hold-overs from early Triassic primitive forms, or degenerate descendents from later types).
Paleorhinus bransomi (Williston 1904) - Pop Agie Formation, Chinle Group, Wyoming; Otis Chalk, Dockum formation, Chinle Group, Howard County, Texas (equatorial northwest Pangea) (= P. scurriensis, based on a partial skull that seems to be a juvenile of P. bransomi)
|
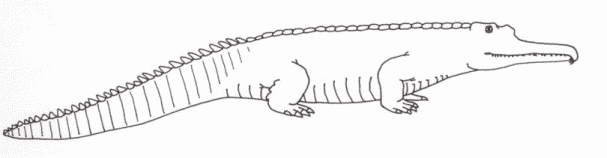 drawing © 2001 Vince R Ward - Prehistoric Pages |
|||
|
Angistorhinus was another of the early phytosaur genera. It was similar to Parasuchus but differed in having the nostrils located further back, towards and above the eyes. This was a more
specialized condition that characterized all phytosaurs above the Parasuchus / Paleorhinus grade. Another
development was that the rear teeth were compressed to blade-like sharp edges, while the front of the snout had cylindrical crushing-type teeth. This was the heterodont (different teeth types) condition, indicating further
specialization over Parasuchus in feeding on large prey. The angistorhinines
were also larger and more heavily built, with crocodile- or alligator-like skulls,
characterized by a shorter, wider, deeper snout. These were huge animals by any standards; the skull length was generally about 120 cm; estimated overall length to 7 or 8 meters, more in Angistorhinus megalodon, a very large, short and heavy-muzzled form sometimes given its own genus (Brachysuchus), and which exceeded in size any living crocodile.
Angistorhinus alticephalus Stovall and Wharton 1936 - Otis Chalk, Dockum formation, Chinle Group, Howard County, Texas. Also reported from Morocco. Skull length 122 cm (type specimen) - shown when alive, left
|
 Illustration by Greg S Paul, from K Padian (ed.), The Beginning of the Age of Dinosaurs. Cambridge Univ. Press. |
|
Saurosuchus galilei,
whose remains have been found throughout the entire stratigraphic range of the Ischigualasto Formation, San Juan Province, Argentina (Southwest Pangea), was an enormous
predator, as big as a full-grown Jurassic theropod dinosaur. Unlike sluggish sprawling present-day reptiles,
Saurosuchus had an erect posture (shown by the elongate ischium and femur) and strong pneumatisation (air-spaces) in the vertebrae [Bonaparte 1970, p.668], which would indicate a metabolically advanced animal. Superior locomotive ability obviously conferred great adaptive advantages, and hence developed among a number of only distantly related archosaurs. (this posture developed in at least three or four distinct clades), and with increased stance came increased metabolic activity. And in this case enabled increase size as well. The skull alone was
up to a metre in length, and the total animal exceeded 6 meters, and must of weighed around three-quarters of a tonne in large specimens.
|
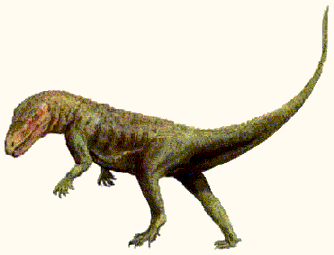 illustration by Steve Kirk, © from Encyclopaedia of Dinosaurs and Prehistoric Animals, Barry Cox, R.J.G.Savage, Brian Gardiner, Dougal Dixon, |
|
Ornithosuchus longidens was a large active terrestrial carnivore, 3 to 4 meters in length, whose remains are known from the Lossiemouth Beds, Elgin, Scotland (Northeast Pangea - Latest Carnian (Adamanian) age). This animal, which was at the top of its local food chain, belonged to the family Ornithosuchidae, a
specialized taxon being known from both Scotland (Lauraisa) and Argentina (Gondwana). The name means "bird crocodile", which seems to nicely describe all the great archosaurs of
Mesozoic, combining as they did features of both crocs and birds, although it is technically only used to describe this group. These were active, fast-moving animals, with fully erect and upright posture, just like dinosaurs, mammals, and
Prestosuchia/Rausichia.
|
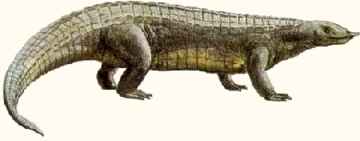 illustration by Steve Kirk, © from Encyclopaedia of Dinosaurs and Prehistoric Animals, Barry Cox, R.J.G.Savage, Brian Gardiner, Dougal Dixon, |
|
The Aetosauridae or Stagonolepidae as they are also known, were an important family of Triassic tetrapods that first appear in the middle or late Carnian. Stagonolepis was a large animal, 2.5 to 2.7 metres in length; I would guess the adult weight at around 200 kg. The body is long and narrow, and the armor lacks the prominent spines of its cousins and contemporaries the desmatosuchines. This was a prominent and widespread animal of the latest Carnian. Two or three species are known. Stagonolepis robertsoni Agassiz (from the Lossiemouth Beds, Elgin), from north-eastern Pangea (Scotland), Stagonolepis wellesi (= Calyptosuschus) (Long & Ballew) (Lower Blue Mesa member, of the Petrified Forest Formation) from equatorial western Pangea (Arizona and New Mexico), although the ranges may have overlapped; and Stagonolepis scagliai (=Aetosauroides) (Casamiquela 1960) from the Ischigualasto Formation, San Juan Province, Argentina. It has been suggested that material from S. scagliai may actually belong to one or other of the other two species, so possibly these may all only be subspecies or variations of the same cosmopolitan form; although cladistic analysis indicates Aetosauroides and Stagonolepis robertsoni are distinct (but related) species. |
 © BBC/ABC from Walking with Dinosaurs |
|
Postosuchus kirkpatricki was a large heavily built animal, some 4 meters in length, known Dockum formation, Chinle Group, Howard Post County, Texas, USA and (Latest Carnian -
Adamanian) the Bull Canyon Formation of east-central New
Mexico (middle Norian); an animal at the top of its food chain. But Postosuchus has a somewhat chequered history hardly befitting its status as top
predator of the Triassic floodplains. It was originally interpreted by its discoverer Sankar Chaterjee as a bipedal
poposaurid and dinosaur ancestor (the poposaurids are now universally considered crocodile uncles), then reinterpreted as a quadrupedal
rauisuchid by Long and Murry 1995. It is furthermore suggested now that the original specimen was based on a confusion of three distinct species, the rauisuchid Postosuchus (most of the skeleton), the poposaurid Lythrosaurus langstoni (part of the hip bones), and the small Chatterjeea elegans (the lightly built forelimbs), an animal of uncertain relationships (either rauisuchid, poposaurid, or member of a distinct family.
|
| Page Back | Unit Home | Page Up | Page Top | Page Next |

Except for material otherwise attributed or quoted, all text on this page is licensed under a
Creative Commons License.
Apart from menu header and other images by MAK or ATW, and public domain gifs, all images on this page are not covered by this
license,
and are copyright their respective owner or publisher.