| Palaeos | Theropoda | |
| Vertebrates | Carcharodontosauridae |
| Page Back | Unit Home | Unit Dendrogram | Unit References | Taxon Index | Page Next |
| Unit Back | Vertebrates Home | Vertebrate Dendrograms | Vertebrate References | Glossary | Unit Next |
DINOSAURIA |--ORNITHISCHIA `--+--SAUROPODOMORPHA | THEROPODA |--Ceratosauria `--Tetanurae |==Megalosauroidea `--Avetheropoda |--Carnosauria | |--Monolophosaurus | `--Allosauroidea | |--Sinraptoridae | `--+--Allosauridae | `--Carcharodontosauria | |--Neovenatoridae | | |--Neovenator | | `--Megaptora | `--Carcharodontosauridae | |--Kelmayisaurus | |--+--Concavenator | | `--Becklespinax | |--Eocarcharia | `--+--Acrocanthosaurus | `--+--Shaochilong | `--Carcharodontosaurinae `==COELUROSAURIA `--AVES |
Contents
Index |
The Carcharodontosaurs constitute a major evolutionary radiation of allosaurs during the Cretaceous period. For the first two thirds of the Cretaceous they had a cosmopolitan duistribution, dominating the large preditor niche and overshadowing the ceratosaurs. During this time, tyrannosaurs were common but limited to the middle preditor niche.
Carcharodontosaurs were a diverse group that included a number of distinct but related lineages - neovenators, acrocanthosaurs, concaventors, carcharodontosaurines, shilantaisaurs, and megaraptors. In a nukmber of thes elines there was a tendency towards gigantism, with lengths in excess of 10 meters being standard. Clearly, like their allosaurine ancestors, these were apex predators that dominated their environments. Of these, the endemic gondwanan carcharodontosaurines were the most impressive.
With the end-Cenomanian extinction event, carcharodontosaur diversity was greatly reduced, and only a single lineage of medium-sized forms, the megaraptors, continued to flourish, mostly in Gondwana, until the end of the Cretaceous. There they co-existed with the uniquely specialised Abelisaurids, which now filled the large predator role in the south, just as tyrannosaurs did in the north.
The recent discovery of this advanced and varied clade of large theropods, previously known only from a few species distributed among the Megalosauridea and Allosauridae, shows once again how paleontology is always making exciting new discoveries about our planet's evolutionary past MAK120304
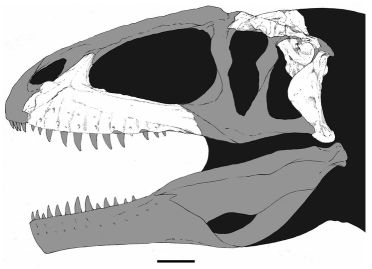
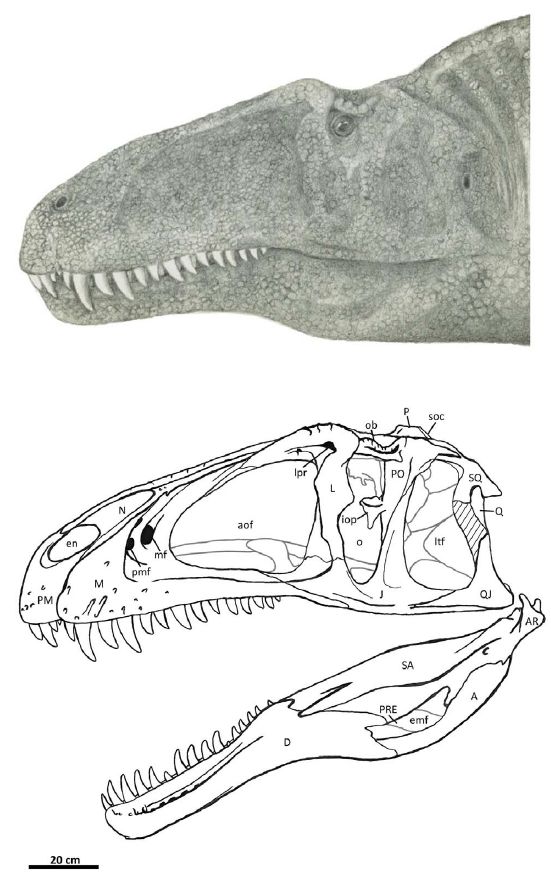 Flesh reconstruction and line drawing of the skull of Acrocanthosaurus atokensis (NCSM 14345) in left lateral view. Hatched lines represent missing bone. A, angular; aof, antorbital fenestra; AR, articular; D, dentary; emf, external mandibular fenestra; iop, intraorbital process of postorbital; J, jugal; L, lacrimal; lpr, lacrimal pneumatic recess; ltf, lateral temporal fenestra; M, maxilla; mf, maxillary fenestra; N, nasal; o, orbit; ob, orbital boss of postorbital; P, parietal; PM, premaxilla; pmf, promaxillary fenestra; PO, postorbital; PRE, prearticular; Q, quadrate; QJ, quadratojugal; SA, surangular; soc, supraoccipital; SQ, squamosal. Image and caption by Eddy & Clarke 2011 fig.2 (doi:10.1371/journal.pone.0017932.g002), Creative Commons Attribution |
Carcharodontosauria Benson et al 2010
Phylogenetic definition: The most inclusive clade comprising Carcharodontosaurus saharicus and Neovenator salerii but not Allosaurus fragilis or Sinraptor dongi (rank free, stem-based) (Benson et al 2010).
Range: Cretaceous of SAm, Afr, Eur, Asia, Aus, N AM
Phylogeny: Allosauroidea : Sinraptoridae + (Allosauridae * : Carcharodontosauridae + Neovenatoridae)
Comments: Characterised by by numerous synapomorphies, for which see the supplementary material by Benson et al 2010. Equivalent to the Carcharodontosauridae of Holtz et al. 2004. All known Cretaceous Carnosaurs and Allosauroids belong here. MAK120226
Neovenatoridae Benson et al 2010 : Neovenator, Chilantaisaurus, Megaraptor
Phylogenetic definition: The most inclusive clade comprising Neovenator salerii but not Carcharodontosaurus saharicus, Allosaurus fragilis or Sinraptor dongi (family rank, stem-based) (Benson et al 2010).
Range: Cretaceous of SAm, Afr, Eur, Asia, Aus, N AM
Phylogeny: Carcharodontosauria : Carcharodontosauridae + * : Neovenator + Megaptora
Comments: Characterised by by numerous synapomorphies, for which see the supplementary material by Benson et al 2010. Equivalent to the Carcharodontosauridae of Holtz et al. 2004. All known Cretaceous Carnosaurs and Allosauroids belong here. MAK120226

Neovenator salerii Hutt, Martill, and Barker, 1996
Horizon: Wealden Clay of Isle of Wight, (Barremian)
Phylogeny: Neovenatoridae : (Chilantaisaurus + Megaptora) + *
Size: length 6 (subadult) to 7.5 meters, weight 0.6 to 1.5 tonnes
Comments: partial skeleton and skull, originally considered an Allosaurid. Later study showed this large animal to be very primitive carcharodontosaurid. It has a very distinctive puffin-like snout. Its discovery, and that of another very primitive contemporary carcharodontosaurian, Concavenator, from the Las Hoyas in Cuenca, Spain, relates the early evolutionary history of this Gondwanan group to the Early Cretaceous of Europe (Ortega et al 2010). Neovenator appears to be a good example of an animal at the evolutionary cross-roads, intermediate or transitional between Allosaurus, primitive Carcharodontosaurs, and Megaptora. This powerful animal would have played an allosaurus-like role in the Early Cretaceous of central Laurasia. MAK010506 revised MAK120302
Image: Neovenator salerii, reconstructed skull, Dinosaur Isle Museum, Isle of Wight, photo by Mo Hassan
 Carcharodontosauridae Stromer 1931 : Giganotosaurus, Carcharodontosaurus > Allosaurus
Carcharodontosauridae Stromer 1931 : Giganotosaurus, Carcharodontosaurus > Allosaurus
Phylogenetic definition: The most inclusive clade comprising Carcharodontosaurus saharicus but not Neovenator salerii Allosaurus fragilis or Sinraptor dongi (family rank, stem-based) (Benson et al 2010).
Range: Early Cretaceous-Late Cretaceous of Eur., Afr., NAm, SAm, & Asia
Phylogeny: Carcharodontosauria : Neovenatoridae + * : (Becklespinax + Concavenator) + Kelmayisaurus + Eocarcharia + (Acrocanthosaurus + (Shaochilong + Carcharodontosaurinae))
Characters: promaxillary foramen of maxilla enlarged to form a fenestra (independently derived in sinraptorids); pneumatic fenestra in maxillary ascending process forming a distinct, open fenestra, unlike in sinraptorids and Ceratosaurus this postorbital contacts lacrimal; postorbital process developed as a small spur (transformed to a large, curving flange in Acrocanthosaurus and more derived carcharodontosaurids); anteroventral end of maxillary paradental groove (defining the dorsal margin of the interdental plates) slants anteroventrally; tall dorsal neural spines more than 1.9 times centrum height. - Benson et al 2010
Kelmayisaurus petrolicus Dong, 1973
Horizon: Lianmugin Formation, of Sichuan, China (Valanginian- Albian)
Phylogeny: Carcharodontosauridae : (Becklespinax + Concavenator) + Eocarcharia + (Acrocanthosaurus + (Shaochilong + Carcharodontosaurinae)) + *
Comments: Kelmayisaurus is one of those fossils where a supposedly anomalous form known only from very fragmentary remains later turns pout to be part of an ordinary lineage of that time. It is based on parts of the upper and lower jaws (maxilla and dentary), both of which have resemblances to both Ceratosaurus and Megalosaurus bucklandii. Originally it seemmd as if, as with Ceratosaurus and Megalosaurs, this was a "living fossil" a persistantly primitive form existing alongside more advanced theropods for millions of years. Restudy shows that this species is more likely a basal carchrodontosaur. . A second species, K. giganticus, based on a vertebral column, will probably turn out to be a sauropod. MAK010506 120302
 Becklespinax holotype BMNH R1828, as illustrated by Richard Owen (A History of British Fossil Reptiles -- Dinosauria Pl. 24) in the 1850s. Owen comments "The extraordinary size and strength of the spines of these anterior dorsal vertebræ, indicate the great force with which the head and jaws of the Megalosaurus must have been used." Via Wikipedia, Public Doamin |
Becklespinax altispinax (Paul,1988)
synonyms: Altispinax dunkeri, Acrocanthosaurus altispinax Paul,1988, Becklespinax altispinax Olshevsky, 1991
Horizon: Wealden Clay of Battle, England (Hauterivian or Barremian)
Phylogeny: Carcharodontosauridae : Kelmayisaurus + Eocarcharia + (Acrocanthosaurus + (Shaochilong + Carcharodontosaurinae)) + (Concavenator + *)
Size: Length 5 to ?8 meters, weight about 1 tonne?
Comments: Known from a set of three tall-spined vertebrae is a genus of large theropod dinosaur based on a type specimen of three tall-spined vertebrae found in 1884 in Sussex, England by the fossil collector Samuel H. Beckles. The Megalosaurus in Crystal Palace, London gets its hump from these vertebrae (Dinosaur Mailing List). (Wikipedia) A partial metatarsus and teeth were also attributed.
The nomenclature of this species is rather convoluted. Becklespinax is based on the high-spined vertebrae that were at one time combined with a Megalosaur tooth species to produce Altispinax dunkeri, which was placed in the family Spinosauridae (on the basis of the vertebrae), being assumed to be a more primitive species in which the vertebrae had not yet extended into a "sail". It was then found that the teeth could not confidently be assigned to the same animal as the postcrania, which were reassigned to a new species of Acrocanthosaurus, another high-spined theropod with similar vertebrae. The species was thus called Acrocanthosaurus altispinax. But further study showed it could not confidently be placed with the genus Acrocanthosaurus either. So it was placed in a new genus and is now known as Becklespinax altispinax. for a while is was placed among the Sinraptoridae, as a smaller version of Yangchuanosaurus, a much earlier animal that likewise had a low fin along the back. The discovery of Concavenator shed new light on this species, both lived around the same time and place, Concavenator lived only slightly (i.e. several millions of years) later), and both have the same set of elongate vertebrae. See also brief comments on Thescelosaurus, and more complete comments at the Theropod Database. At the time of writing, the two have not been described in a peer reviewed paper. MAK010506 revised 120226
Concavenator corcovatus Ortega, Escaso and Sanz 2010 (possibly = Becklespinax corcovatus (Ortega, Escaso and Sanz) ?)
Horizon: Calizas de La Hue´rguina Formation, Late Barremian (Early Cretaceous); Las Hoyas, Cuenca, Spain
Phylogeny: Carcharodontosauridae : Kelmayisaurus + Eocarcharia + (Acrocanthosaurus + (Shaochilong + Carcharodontosaurinae)) + (Becklespinax + *)
Size: about 6 meters long
 Concavenator corcovatus using its hump as a thermoregulatory device, absorbing sunlight in the warmth of a morning sunrise. Artwork by Emily Willoughby, via Wikipedia, Creative Commons Attribution |
Comments: Concavenator corcovatus, the "hump backed hunter from Cuenca", is a recently discovered dinosaur from the Las Hoyas lagerstatte, discovered by José Luis Sanz, Francisco Ortega and Fernando Escaso from the Autonomous University of Madrid and the National University of Distance Learning.
Apart from Neovenator, Concavenator is the most primitive known carcharodontosaurid. It is the most complete individual of a carcharodontosaurian theropod, and the first one in which direct and indirect evidence of integumentary structures is reported. This animal is remarkable for the presence of two extremely tall vertebrae in front of the hips formed a tall but narrow and pointed crest on its back. The function of such a crest is not clear; it could be like the head-crests used in intra-species visual displays, or it could also be a thermal regulator (see illustratuion at right), rather the fin or sail on the back of edaphosaur and sphenacodontid pelycosaurs. Similar elongate vertebrae are found in spinosaurs (although they have a longer "sail")
Another curious feature is that the forelimb (ulna) shows evidence of what may be quill knobs or homologous structures, an anatomical feature so far known only in animals with large, quilled feathers on the forelimb. If this is the correct explanation, it shows that feathers appared much earlier in theropod evolution than previously thought (at the avetheropod rather than the maniraptoran grade). Although it is less surprising in view of the presence of tubular quill-like structures in ornithischian dinosaurs such as the heterodontosaurid Tianyulong and the basal ceratopsian Psittacosaurus (Ortega et al 2010, p.205). Darren Naish of tetrapod zoology is sceptical of this interpretation, pointing out that the bumps were unusually far up the arm and irregularly spaced for quill knobs. His alternative explanation is that such structures are found along intermuscular lines, acting as tendon attachment points for example. (Naish 2010b) modified from Wikipedia MAK120302
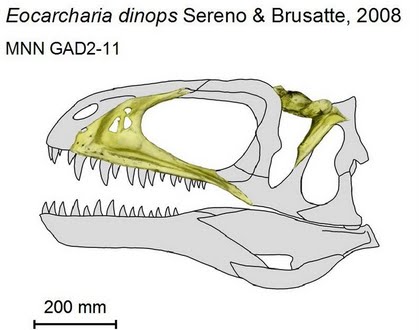
Eocarcharia dinops Sereno & Brusatte, 2008 (possibly = Acrocanthosaurus dinops (Sereno & Brusatte) ?)
Horizon: "Gadoufaoua" on the western edge of the Ténéré Desert, Niger; Elrhaz Formation (Aptian-Albian).
Phylogeny: Carcharodontosauridae : (Becklespinax + Concavenator) + Kelmayisaurus + ((Shaochilong + Carcharodontosaurinae) + Acrocanthosaurus) + *
Comments: Eocarcharia (meaning "dawn shark", in reference to its basal status in the "shark-toothed" dinosaurs) is a carcharodontosaurid from the early part of the Middle Cretaceous. its fossil remains were discovered in the Sahara desert (where in places dinosaur bones are just appearing out of the sand dunes) in 2000 on an expedition led by University of Chicago paleontologist Paul Sereno (Wikipedia). It is represented by several cranial bones (fragments of skull & jaws) and isolated teeth. The species name dinops is from the Greek "fierce-eyed", in reference to the massive ornamented brow above the orbit.
This basal carcharodontosaurid is similar to and contemporary with Acrocanthosaurus. (Sereno & Brusatte 2008). The two are sometimes grouped in a single clade, so it is possible they are congeneric, although Paul 2010, who is (like the present author (MAK) a lumper when it comes to species and genera, does not do so here. in any case, the known remains of Eocarcharia are so fragmentary that is difficult to be sure of its exact phylogenetic plaxcement, beyond basal carcharodontosaurid. The presence of a close relative of the North American Acrocanthosaurus on Africa suggests that carcharodontosaurids had already achieved a trans-Tethyan distribution by the mid Cretaceous. (Sereno & Brusatte 2008). This animal was a contemporary of the abelisaurid, Kryptops. The co-exstence of several totally different lineages of large theropods was a common occurance from the Late Jurassic through to the Middle Cretaceous. MAK120226
Image: type specimen of Eocarcharia dinops; illustration by Norbu Tamura, following Sereno & Brusatte 2008; from Palaeocritti
 Acrocanthosaurus:
Stovall & Langston, 1950. A. atokensis Stovall & Langston,
1950.
Acrocanthosaurus:
Stovall & Langston, 1950. A. atokensis Stovall & Langston,
1950.
Range: Early Cretaceous (Aptian-Albian) of North America (Oklahoma)
Phylogeny: Carcharodontosauridae : (Becklespinax + Concavenator) + Kelmayisaurus + Eocarcharia + (Shaochilong + Carcharodontosaurinae) + *)
Characters: 6+m [CC00]; Dermal skull: skull 129cm long [CC00]; facial horns
absent [CC00]; premaxillae meet at acute angle, forming sharp, narrow anterior margin [CC00];
subnarial foramen between premaxilla & maxilla [CC00]; antorbital fenestra
very large & triangular [CC00]; antorbital fenestra with overhanging
anterioventral rim, covering small promaxillary fenestra [CC00]; nares small
& elongate [CC00]; naris formed by premaxilla & nasal, excluding maxilla
[CC00]; maxilla with 2 accessory fenestra [CC00]; maxilla taller than long
[CC00]; maxilla, anterior margin slopes posterodorsally [CC00]; maxilla forms
most of anterior margin of antorbital fenestra [CC00]; maxilla
rugose only
anteriorly & near tooth row [CC00]; maxilla posterodorsal process passes
between nasal and antorbital fenestra, bifurcating distally to embrace
anteroventral process of lacrimal.[CC00]; maxillary fenestra between naris &
antorbital fenestra [CC00]; nasal flat, narrow, 4x length of frontal [CC00];
nasal with groove for maxilla articulation, allowing some lateral rotation of
maxilla [CC00]; nasal overlaps lacrimal & frontal [CC00]; lacrimal forming
low ridge anterior to orbit (but not horn as in Sinraptor) [CC00]; lacrimal contacts postorbital
under lappet of prefrontal [CC00]; "the posterior margin of the ventral
(preorbital) process of the lacrimal has a double concavity separated by a short
process (for attachment of the ligamentum suborbitale) that 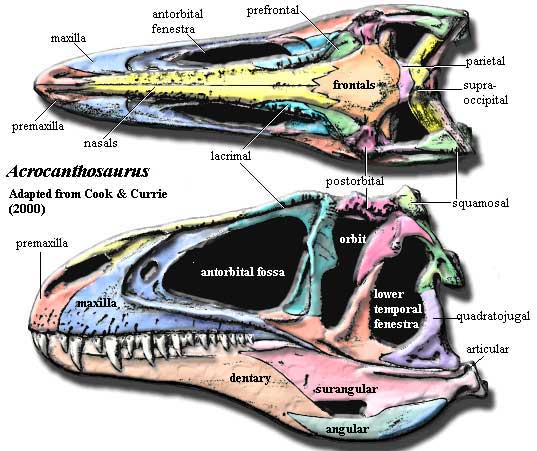 marks the
anteroventral extent of the eyeball itself (diameter was about 11 cm)"
[CC00]; nasal & lacrimal with
straight, slightly rugose ridge separating dorsal & lateral exposures
[CC00]; prefrontal large & triangular [CC00]; prefrontal contacts
postorbital & may participate in orbit, excluding frontal [CC00]; frontals
"more or less" fused, as is frontal-parietal suture [CC00]; skull roof narrow [CC00];
parietals perpendicular to vertical occiput [CC00]; sagittal crest absent
[CC00]; jugal participates in antorbital fenestra [CC00]; jugal overlaps
lacrimal so that lacrimal-maxilla contact absent [CC00]; jugal form
posteroventral margin of orbit [CC00]; jugal, postorbital process huge &
triangular [CC00]; orbit key-hole shaped & narrow [CC00];
cheek flat, vertical, at sharp angle to skull roof [CC00]; postorbital with
thick, rugose rim [CC00]; postorbital, intertemporal ramus short and almost
vertical [CC00]; postorbital forming wide postorbital bar [CC00]; lower
temporal
fenestra piriform & bordered dorsally by short
intertemporal bar [CC00];
jugal & quadratojugal moderately invade lateral temporal fenestra [CC00];
quadratojugal overlaps quadrate, curving onto occipital surface [CC00];
squamosal with deep quadrate cotylus, probably exposing head of quadrate in
lateral view [CC00]; "Posteroventrally, the squamosal extended along the
dorsal surface of the paroccipital
process, covering up the ventrolateral
process of the parietal and wrapping around onto the anterodorsal surface of the
distal end of the paroccipital process. The squamosal was inclined in such a
manner that it would have roofed over the posterior temporal musculature"
[CC00: 217]; Occiput: supraoccipital
forms double boss behind low
nuchal crest [CC00]; supraoccipital with strong
midline ridge [CC00]; paroccipital process broad & shallow, turning
ventrally at lateral ends, without distal posterior projection [CC00];
occipital condyle circular, not flattened [CC00]; Mandible: jaw with
large external mandibular fenestra [CC00]; dentary long and low, laterally
smooth with large mental foramina [CC00]; dentary with ventral process under
symphysis [CC00]; angular with thickened ventral rim [CC00]; angular having
sliding contact with dentary [CC00]; retroarticular
process oriented posteromedially [CC00]; supradentary
present, unknown if fused to coronoid [CC00]; Dentition: premaxilla with 4 teeth, maxilla with 15 teeth (dentary count
unknown) [CC00]; teeth without wrinkled enamel [CC00]; premaxillary teeth
blade-like, not D-shaped [CC00]; Axial: body high and narrow
[CC00]; neural spines >2.5x length of
centra [CC00]; precaudal centra strongly excavated with complex pleurocoels [CC00]; cervical neural spines with triangular anterior processes inserting into
depressions under overhanging processes on preceding neural spines [CC00]; mid-
and distal caudals with posteriorly placed neural spines [CC00]; mid-caudal vertebrae with accessory transverse processes
[CC00]; Appendicular: arm short (81% femur length) [CC00]; scapula
long slender, gently curved [CC00]; deltopectoral
crest large, quadralateral [C00].
marks the
anteroventral extent of the eyeball itself (diameter was about 11 cm)"
[CC00]; nasal & lacrimal with
straight, slightly rugose ridge separating dorsal & lateral exposures
[CC00]; prefrontal large & triangular [CC00]; prefrontal contacts
postorbital & may participate in orbit, excluding frontal [CC00]; frontals
"more or less" fused, as is frontal-parietal suture [CC00]; skull roof narrow [CC00];
parietals perpendicular to vertical occiput [CC00]; sagittal crest absent
[CC00]; jugal participates in antorbital fenestra [CC00]; jugal overlaps
lacrimal so that lacrimal-maxilla contact absent [CC00]; jugal form
posteroventral margin of orbit [CC00]; jugal, postorbital process huge &
triangular [CC00]; orbit key-hole shaped & narrow [CC00];
cheek flat, vertical, at sharp angle to skull roof [CC00]; postorbital with
thick, rugose rim [CC00]; postorbital, intertemporal ramus short and almost
vertical [CC00]; postorbital forming wide postorbital bar [CC00]; lower
temporal
fenestra piriform & bordered dorsally by short
intertemporal bar [CC00];
jugal & quadratojugal moderately invade lateral temporal fenestra [CC00];
quadratojugal overlaps quadrate, curving onto occipital surface [CC00];
squamosal with deep quadrate cotylus, probably exposing head of quadrate in
lateral view [CC00]; "Posteroventrally, the squamosal extended along the
dorsal surface of the paroccipital
process, covering up the ventrolateral
process of the parietal and wrapping around onto the anterodorsal surface of the
distal end of the paroccipital process. The squamosal was inclined in such a
manner that it would have roofed over the posterior temporal musculature"
[CC00: 217]; Occiput: supraoccipital
forms double boss behind low
nuchal crest [CC00]; supraoccipital with strong
midline ridge [CC00]; paroccipital process broad & shallow, turning
ventrally at lateral ends, without distal posterior projection [CC00];
occipital condyle circular, not flattened [CC00]; Mandible: jaw with
large external mandibular fenestra [CC00]; dentary long and low, laterally
smooth with large mental foramina [CC00]; dentary with ventral process under
symphysis [CC00]; angular with thickened ventral rim [CC00]; angular having
sliding contact with dentary [CC00]; retroarticular
process oriented posteromedially [CC00]; supradentary
present, unknown if fused to coronoid [CC00]; Dentition: premaxilla with 4 teeth, maxilla with 15 teeth (dentary count
unknown) [CC00]; teeth without wrinkled enamel [CC00]; premaxillary teeth
blade-like, not D-shaped [CC00]; Axial: body high and narrow
[CC00]; neural spines >2.5x length of
centra [CC00]; precaudal centra strongly excavated with complex pleurocoels [CC00]; cervical neural spines with triangular anterior processes inserting into
depressions under overhanging processes on preceding neural spines [CC00]; mid-
and distal caudals with posteriorly placed neural spines [CC00]; mid-caudal vertebrae with accessory transverse processes
[CC00]; Appendicular: arm short (81% femur length) [CC00]; scapula
long slender, gently curved [CC00]; deltopectoral
crest large, quadralateral [C00].
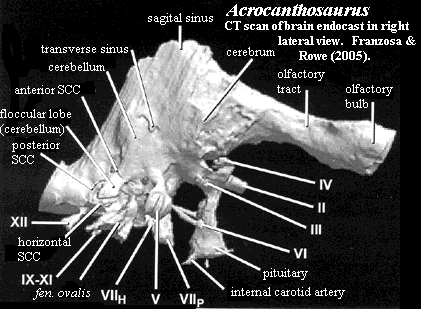
References: Currie & Carpenter (2000) [CC00]; Franzosa & Rowe (2005) [FR05]. ATW060217.
Size: length 8 to 12 meters, weight upto 3 to 4.4 tonnes
Comments: A huge preditor, filling the same ecological role in the north as the Carcharodontosaurines did in the south, Acrocanthosaurus is distinguished by tall vertebral spines giving it a metricanthosaur-like appearance. The basal carcharodontosaurid Becklespinax has a similar but taller fin. Acrocanthosaurus, or an animal very much like it, made some of the famous Paluxy River dinosaur tracks in Texas, including a sequence where an individual appears to pursue a large brachiosaurid (probably Pleurocoelus), which also left its geeat basin-sized prints in the mud. (silly trivia note: creationists argue that some of the Pleurocoelus fore-foot prints are actually "man tracks" made by giant humans). This indicates that this Tyrannosaurid-sized carnivore was not afraid to take on even the biggest sauropods in its environment. Represents a transitional form between allosaurids and primitive charcharodontosaurids and carcharodontosaurines proper. This was the last-known large Allosaurian in North America (Western Laurasia), apart from a relict lineage (Labocania) in the very South-West of the continent. The Acrocanthosaurs disappear at the end of the mid Cretaceous, perhaps victims of the Cenomanian mass-extinction. The Tyrannosaurids take their place in the north. In the south, the Carcharodontosaurines are replaced by Abelisaurs. MAK010506 revised MAK120226
Use of the forelimb in handling prey: Unlike other highly specialised large theropods like Abelisaurs and Tyrannosaurs, Acrocanthosaurus would have used its small but functional forearms to secure prey. The range of motion in the forelimb of Acrocanthosaurus is similar to that of Herrerasaurus and Dilophosaurus, and greater than that of Tyrannosaurus. The limited forward range of motion of the forelimb indicates that Acrocanthosaurus captured prey with its jaws. The forelimb could grasp prey that had been moved by the jaws to beneath the chest. Struggling prey would have been impaled upon the strongly flexed first ungual. Acrocanthosaurus exhibits a greater manual range of motion than ornithomimid and deinonychosaurian coelurosaurs, but less at the shoulder and elbow. Coelurosaurian theropods exhibit reduced digital flexion and hyper-extension, which suggests a change in the use of the hand. Senter & Robins 2005) MAK120307
Note: usually placed in the Carcharodontosauridae, less often in the Allosauridae. But see, Currie & Carpenter (2000).
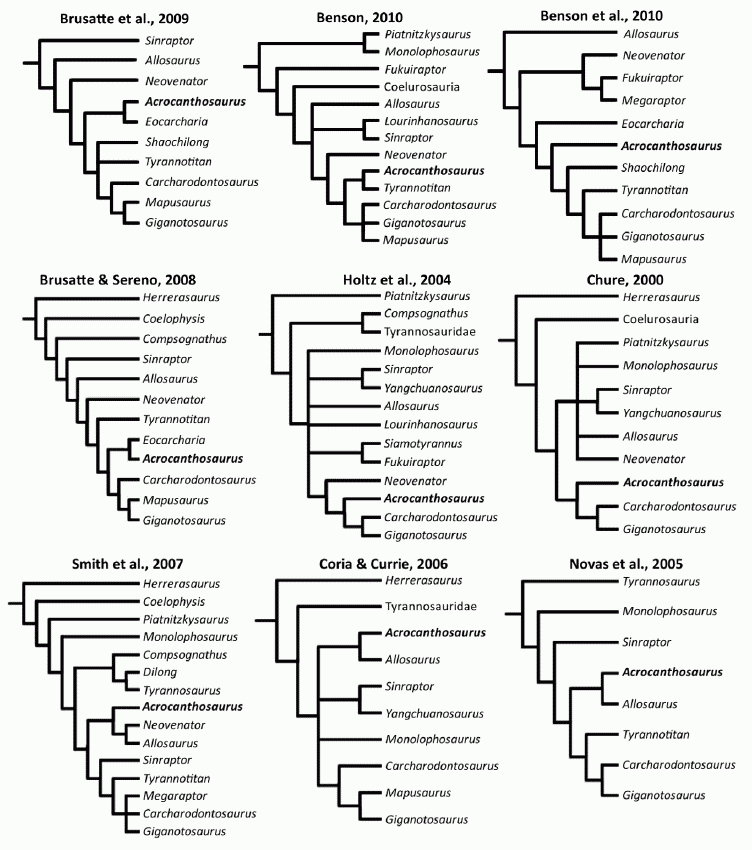 Nine phylogenetic hypotheses, showing various interpretations of Acrocanthosaurus in the Allosauroidea. Analyses along the top two rows recover Acrocanthosaurus atokensis as a member of Carcharodontosauridae, whereas the bottom row represents those that place Acrocanthosaurus atokensis as close relative and/or sister taxon to Allosaurus. Image and some of caption by Eddy & Clarke 2011 fig.33 (doi:10.1371/journal.pone.0017932.g033), Creative Commons Attribution |
 Reconstruction of the skull of Shaochilong maortuensis, from Brusatte et al 2010b, based on IVPP V2885. Known parts in white. Scale = 5 cm. Illustration by Brett Booth Original url |
Shaochilong maortuensis Brusatte, Benson, Chure, X. Xu, Sullivan, and Hone, 2009
Synonym: Chilantaisaurus maortuensis Hu, 1964, Alashansaurus
Horizon: Ulansuhai Formation of Outer Mongolia (Turonian)
Phylogeny: Carcharodontosauridae : (Becklespinax + Concavenator) + Kelmayisaurus + Eocarcharia + (Acrocanthosaurus + (Carcharodontosaurinae + *))
Size: based on the length of the maxillary tooth row, estimated length of 5 to 6 metres (16 to 20 ft). Based on partial femur, weight approximately 500 kilograms
Comments: Known from skull fragments and some vertebrae, and it seems some limb bones, Shaochilong (meaning "shark toothed dragon") is unusual both in its northern distribution and late straigraphic range. Originally identified as a carnosaur of the genus Chilantaisaurus and then a primitive coelurosaur (Wikipedia), a re-description by Brusatte and coworkers in 2009 found that it was in fact a carcharodontosaurid, the first recognized from Asia. Phylogenetic analysis performed by indicate that Shaochilong is deeply nested within the carchorodontosaurids, and appears to be more closely related to the classic Gondwanan carcharodontosaurids (Tyrannotitan, Carcharodontosaurus, Mapusaurus, Giganotosaurus) than the Laurasian ones (such as Neovenator and Acrocanthosaurus). It shows that allosaurs rather than tyrannosaurs were still the dominant group of large-bodied theropods in Laurasian as late as the early Late Cretaceous. Compared to other taxa however it was a small bodied and short-snouted representative of the group. The dominance of the tyrannosaurids in Asiamerica was therefore confined to the very end of the Cretaceous. (Brusatte et al 2009, Brusatte et al 2010b)
Shaochilong maortuensis may or may not be related to Labocania anomala Molnar, 1974 of the Late Campanian El Gallo Formation, Baja California, Mexico, this large but very poorly known animal (estimated length about 7.5 m; weight about 1.2 tons) has been variously interpreted as a tyrannosaur-like allosaurid (Paul 1988) a possible tyrannosauroid (Holtz2004), and an avemetatarsalian, it has also been related to Shaochilong Chure (2000) . Mickey Mortimer suggests carcharodontosaurid related to Shaochilong. As East Asia and Western North America were closely connected during the late Cretaceous, this makes sense from a paleogeographic as well as stratigraphic perspective. If so, two distinct lineages of carcharodontosaurs survived till the end of the Cretaceous, showing that late Cretaceous large theropod diversity was a lot greater than previously realised. This also means that allosaurs were independently evolving tyrannosaur characteristics. Alternatively, and rather more likely given the stratographic and geographic location and morphological characteristics, this could simply be another basal tyrannosaur. In this case, no carcharodontosaurids survived to end of the Mesozoic. MAK120302
| Page Back | Unit Home | Page Top | Page Next |
checked ATW050619, page MAK120302