Some Fossils
| Palaeos: | Sarcopterygii | |
| The Vertebrates | Ganthostome Orbits -- Some Fossils |
| Page Back | Unit Home | Unit Dendrogram | Unit References | Taxon Index | Page Next |
| Unit Back | Vertebrates Home | Vertebrate Dendrograms | Vertebrate References | Glossary | Unit Next |
|
Abbreviated Dendrogram
Teleostomi |--Neopterygii `--Sarcopterygii |--+--Onychodontiformes | `--Actinistia `--Rhipidistia |--Dipnomorpha `--+--Rhizodontiformes `--Osteolepiformes |--Tristichopteridae `--Elpistostegalia |--Panderichthys `--Tetrapoda |
Contents
Overview |
 Orbiting
the Gnathostomes: Some Fossils
Orbiting
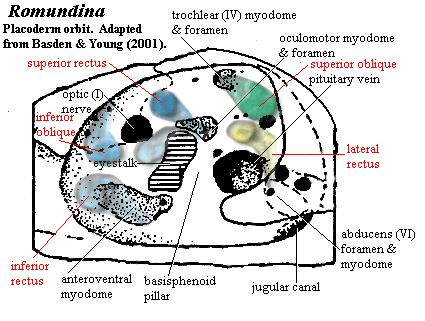
the Gnathostomes: Some FossilsNow we are in a better position to look at some real fossils from the region of interest. The reason we must is that we may not entirely see eye to eye with some recent interpretations in this area. The first figure is the (perhaps basal) placoderm, Romundina. For the most part, this is entirely in agreement with our model. However, note the position of the anteroventral myodome, which we have given a carefully neutral name. Basden & Young (2001), building on the work of Young (1986) [1], call this the "ventral myodome" and assign the enclosed foramen to the ophthalmic artery. Note, however, that it fades out anteriorly, suggesting that it houses the inferior rectus. So, we may expect that a branch of the oculomotor nerve is somewhere in the picture. However, it then becomes difficult to place the inferior oblique, which ought to oppose it. The problem is overcome if we place the inferior oblique along the anterodorsal edge of the orbit, and assume it is ennervated through one of the three small foramina in thast region. This results in a pattern which appears peculiar only because the entire complement of muscles is rotated clockwise with respect to the shark.
The next image is the orbit of Lingulalaepis, the very basal actinopterygian whose braincase was recently described by Basden & Young (2001). We have then reconstructed the extrinsic eye muscles in accordance with our model. For the most part, everything works out relatively nicely. However, there are a couple of problem areas which require comment.
The most significant difficulty is the anteroventral myodome mentioned
above. In our reconstruction, we have assigned it to the 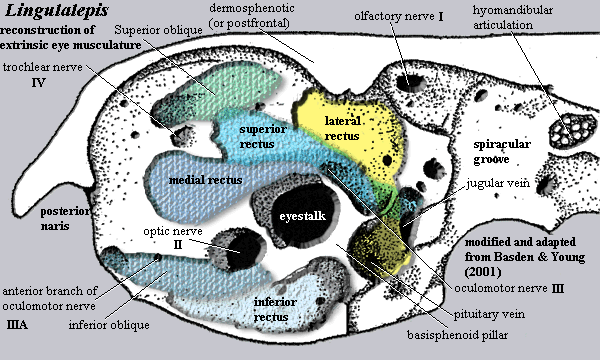 inferior oblique
muscle. A nearby foramen, apparently just hanging around waiting to get
into trouble, has been assigned to productive work as an anterior branch
of the oculomotor nerve. As far as we are able to make out, Basden &
Young assign this myodome to nothing. The depression also contains a small foramen (almost invisible in the image)
which could also admit an oculomotor branch. Basden & Young label it
"ventral nasobasal canal" a structure homologous -- or analogous
-- to the
nasolacrimal duct of sarcopterygians. However, if it is invisible in anterior view, as
their figure 7 (not shown) suggests, then this foramen might, alternatively, be a passage for the
oculomotor nerve approaching from the posterior of the orbit and closely
integrated with the medial surface of the orbital wall.
inferior oblique
muscle. A nearby foramen, apparently just hanging around waiting to get
into trouble, has been assigned to productive work as an anterior branch
of the oculomotor nerve. As far as we are able to make out, Basden &
Young assign this myodome to nothing. The depression also contains a small foramen (almost invisible in the image)
which could also admit an oculomotor branch. Basden & Young label it
"ventral nasobasal canal" a structure homologous -- or analogous
-- to the
nasolacrimal duct of sarcopterygians. However, if it is invisible in anterior view, as
their figure 7 (not shown) suggests, then this foramen might, alternatively, be a passage for the
oculomotor nerve approaching from the posterior of the orbit and closely
integrated with the medial surface of the orbital wall.
This is at least moderately consistent with the presence of an anterior myodome in other basal actinopterygians, such as Mimia (below). To be sure, the anterior myodome in Mimia is a double structure and is located anterodorsally. However, it may well represent, for example, a near-convergence of the origins of both oblique muscles.
By contrast, Basden & Young tentatively assign the inferior oblique muscle to the ventral myodome where we have placed the inferior rectus. Their discussion of the superior, medial and inferior rectus does not allow one to place those muscles with any confidence. However, it is difficult to create the necessary muscle oppositions if the inferior rectus and oblique originate in more or less the same place.
The problem may stem from Basden & Young's use of
placoderms as the model model. As we have 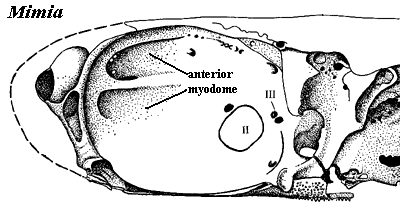 seen,
the placoderm orbit is unique because of the approximately 45° clockwise
rotation of the muscle origins. We prefer a reconstruction which resembles the case of sharks
and coelacanths, is not completely at odds with the apparent geometry in
placoderms, makes mechanical sense, and keeps appropriate symmetry between the
dorsal and ventral halves of the orbit -- at least until the next basal
gnathostome braincase is described.
seen,
the placoderm orbit is unique because of the approximately 45° clockwise
rotation of the muscle origins. We prefer a reconstruction which resembles the case of sharks
and coelacanths, is not completely at odds with the apparent geometry in
placoderms, makes mechanical sense, and keeps appropriate symmetry between the
dorsal and ventral halves of the orbit -- at least until the next basal
gnathostome braincase is described.
Unfortunately, in both actinopterygians and sarcopterygians the story becomes very hard to follow past a certain point. The loss of the eyestalk altered the mechanical properties of the eyeball to rotation and seems to have resulted in a major shift and reduction in the extrinsic eye muscles. Nevertheless, we may attempt to apply our crude model to some basal sarcopterygians in the following pages. ATW040416.
[1] Young, GC (1986), The relationships of placoderm fishes. Zool. J. Linn. Soc. 88: 1-57.
| Page Back | Unit Home | Page Top | Page Next |
checked ATW041103