| Palaeos: | Euteleostei | |
| The Vertebrates | Overview |
| Page Back | Unit Home | Unit References | Unit Dendrogram | Taxon Index | Page Next |
| Unit Back | Vertebrates Home | Vertebrate Dendrograms | Vertebrate References | Glossary | Unit Next |
|
Abbreviated Dendrogram
Teleostei | Euteleostei |--Ostariophysi | |--Anotophysi | `--Otophysi | |--Cypriniformes | `--Siluriphysi `--Neognathi |--+--Salmoniformes | `--Esociformes `--+--Argentiniformes `--Neoteleostei |--Stenopterygii `--Eurypterygii |--Cyclosquamata `--Ctenosquamata |--Scopelomorpha `--Acanthomorpha |
Contents
Overview |
As suggested in the taxon summary, there is considerable doubt whether the Euteleosts are a clade. The uniques characters of the Euteleosts, such as seasonal tubercles on breeding males, are all of a type which are usually untestable on the fossil record. Since the basal Teleost lineage appeared some time in the Triassic, we may never know whether or not basal Teleosts shared these characters. As a result, this taxon has fallen out of favor in fish systematics.
This is probably just as well, although it hard on the student of taxonomy. There are thousands of teleost species. Teleosts are, in fact, the most gloriously successful group of vertebrates the world has ever seen. One would like to have some useful higher level taxa to organize the spectacular diversity of this taxon. However, the fish have been unaccountably uncooperative. They persist in diversifying to suit their individual needs with no thought at all to the difficulties this imposes on central taxonomic planning. Undoubtedly, some committee is hard at work in Brussels or Washington to establish a more socially responsible scheme of speciation. However, pending these developments, we deal, perforce, with fish as they are.
Teleosts are frequently identified by their jaw structure. In
fact, the principle acheivement of the teleosts is their terminal, highly
articulated mouth in which the posterior end of the upper jaw rotates forward on
an anterior pivot as the lower jaw drops. This creates a large round
opening which projects forward and sucks food items into the mouth. The precise
mechanics and the bones involved differ somewhat between species, and it appears
that the mechanism has evolved convergently a number of times. As a result,
anatomists searching for consistent trends have often looked to the other end of
the fish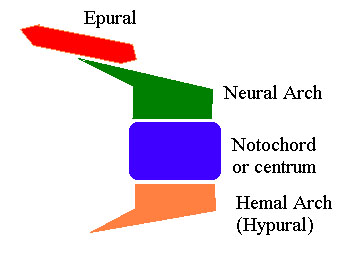 .
.
The Teleost tail is made up of individual modified vertebral units which are illustrated schematically in the figure (in right lateral view). Not all of these elements will necessarilly be present in any one segment. In addition, a number of dermal elements may be attached notably fin rays:
(a) The most common (and primitive) dermal elements are fin rays which extend from the distal ends of the hypural spines and form the framework of the lower fin lobe.
(b) Distally, the central elements disappear and the line of free hypurals curves dorsally to meet the line of free epurals (if present) or of neural arches. At the intersection, the hypural spines are now directed caudally, or even dorso-caudally. These terminal hypurals frequently develop a large number of fin rays on their dorsal surfaces which become the rays of the upper caudal fin lobe.
Any number of varriations are possible and may be diagnostic. For example, the neural spines of vertebral units anterior to the tail may become very elongated and participate in the upper lobe. Frequently some of the more distal elements (hypurals, spines, or even epurals) fuse or become dorsoventrally expanded into larger, more-or-less triangular elements (i.e., with the base of the triangle directed outward). Scales adjacent to the tail may elongate and participate in the structure of the tail as caudal scutes or fulcral scales.
In the earliest fish, the notocord penetrated to the very end of the tail. Even where the tail was externally bi-lobed and symmatrical, the notocord extended to the end of one of the lobes, normally the upper lobe in gnathostomes. However, the tendency has been for the vertebral column to retreat from the tail.
The received lesson from all this is that teleosts gradually perfected the symmetrical, homocercal tail (i.e. a tail with both upper and lower lobes having a similar external morphology and internal structure) which, with the general loss of heavy armored scales, allowed them to speed away from predators and to become fruitful and multiply. The feat was accomplished by evolving methods for stabilizing the tail in both lobes without requiring cordal or vertebral support in either lobe, thus making the modern teleost tail a light, strong but flexible control device despite the absence of either axial bone or muscle.
There are several difficulties with this view of teleost tail development. The development times may be too long. Both mammals (whales) and reptiles (ichthyosaurs) returning to the sea developed quite workable homocercal tails in very short order. By contrast, chondrichthyans, a group which has included dedicated fast swimmers since at least the Devonian, seem to be quite happy with an assymmetrical tail. In fact, the very diversity of teleost tail structure suggests that tail structure may not be not a particularly critical factor and that a change in tail structure is not a process which ought to take over 100 My.
This is not to say that the conventional fish tale is wrong. However, it may have a rather more complex plot. In those few cases where a particular, definable genetic or morphological change is clearly advantageous, evolution seems to proceed very quickly. The fact that many, comparatively straightforward, pieces of engineering such as the teleost tail seem to take a very long time may have a number of interesting explanations. Three posibilities are discussed here.
First, the change may not be as clear as we believe. That is, the structure of the tail cannot be isolated from everything else going on in the organism. For example, changes in tail structure may affect, and be affected by the nature of the axial skeleton (e.g. notocord vs. vertebrae), the axial musculature, the neurological changes necessary to rewire a swimming style, ecological factors favoring speed, dietary factors controlling relative growth of tissues, sexual selection, and so on. If the parts are sufficiently inter-related, we cannot make sensible statements about selection in a single module such as the tail, and the complex interrelated changes which take place may be slow.
Second, the selective advantage may not be clear. Free lunches are rare. A favorable change in one feature commits resources in a way which causes may cause unfavorable changes elsewhere. The emerging story of the dinosaurs is also a cautionary tale. We should not casually assume that earler forms were inferior in adaptation to their particular environment. Certainly, over 100 My, the chance that the environment has changed in a relevant way is at least as high as the chance that the tail engineering has actually "improved." Perhaps the starting assumption should be that species are near equilibrium with respect to their environment and optimally adapted for their body plan and niche within it.
Third, for reasons which are not completely understood, individual species tend to be very stable in overall form, even where there is substantial genetic diversity within the population (e.g. the extremely conservative horseshoe crab, Limulus). If the stabilizing tendency, whatever it may be, is sufficiently strong, it will defeat both selection and genetic drift, the primary engines of change. If so, then species will be far from adaptational equilibrium and the rate-limiting step in evolution becomes speciation. Since speciation requires the creation of a sexually isolated group (usually by physical separation or allopatry), it tends to be a relatively rare event. Consequent change at the family level is thus slower, and tends to procede in jumps. This alternative is, as readers will recognize, merely a restatement of Gould & Eldridge's punctuated equilibrium model.
To some degree, all three processes may be taking place. Possibilities 2&3 are, however, mutually inconsistent in a testable way, since 2 assumes processes close to adaptational equilibrium and 3 violates this asumption. However 1 cautions us that the tests may not be all that they seem. Just as it may not be possible to view a tail in isolation, it may not be possible to view even an organism in isolation. But the level(s) at which evolution operates -- gene, anatomical module, organism, species, ecosystem -- is beyond the scope of an introduction already too long.
| Page Back | Unit Home | Page Top | Page Next |
checked ATW030905