| Palaeos: | Conodonta | |
| The Vertebrates | Overview (2) |
| Page Back | Unit Home | Unit Dendrogram | Unit References | Taxon Index | Page Next |
| Unit Back | Vertebrates Home | Vertebrate Dendrograms | Vertebrate References | Glossary | Unit Next |
|
Abbreviated Dendrogram
Vertebrata |--+--Pteraspidomorphi | `--Gnathostomata Conodonta |--Paraconodontida `--Euconodonta |--Proconodontidae `--+--Protopanderodontida `--Prioniodontida |--Balognathidae `--+--Prioniodinida `--Ozarkodinida |
Contents
Index |
 On
a previous page, we indicated doubt -- in fact outright rejection -- of the
conodont eye, as interpreted by Aldridge, Donoghue and others. On this
page, we present the defense of our position, together with an alternative
interpretation of the conodont head based on the same, critical IGSE 13821
specimen.
On
a previous page, we indicated doubt -- in fact outright rejection -- of the
conodont eye, as interpreted by Aldridge, Donoghue and others. On this
page, we present the defense of our position, together with an alternative
interpretation of the conodont head based on the same, critical IGSE 13821
specimen.
For reasons much too obvious to discuss, we will refer to the currently floated hypothesis as the "Crawling Eye" theory of conodont structure. In the 1958 movie of this name, a strange radioactive cloud settles on top of a remote mountain. After interminable mood music, and what passes for character development in this genre, an evil alien emerges from the cloud. By some bizarre quirk of interstellar homoplasy, it looks like a giant human eyeball with tentacles. But, supposing this to be true, what are the chances that, over the parsecs and megayears, we can take this similarity at, as it were, face value?
For the same reasons, we must sternly resist the temptation to assume that every large, bilateral, hemispherical structure -- even on an Earthly chordate -- is necessarily an eye. We will discuss a number of factors which make the Crawling Eye interpretation of IGSE 13821 unlikely; following which, we will present an alternative model which seems more consistent with the known information about conodonts.
Before braving the perils of morphology and differential tissue preservation, let us consider the difficulties of the Crawling Eye model a priori. It is a relatively simple matter to make a short list.
1. Not to belabor the obvious, but that's a hell of a big eye for a little beastie like a conodont. What happens to all that visual information? Where is the optic tectum or forebrain necessary to process the images? Heavy reliance on visual information is neurologically expensive and requires a great deal of cerebral overhead -- quite literally -- to make proper use of the hypertrophied sensory organ.
2. The orbits bear a close relationship to the adenohypophysis from the point of view of genetic regulation. See, e.g., The Basisphenoid. In most basal vertebrates that association is also physical, so that the sella turcica or its predecessor lies in close proximity to the orbits or to an interorbital bar. While this is not always the case, the absence of anything similar here is suspicious.
3. The "accessory lobes," two round, dark patches posteromedial to the "eyes" are supposed to be otic capsules. This should inspire a brief, but urgent, reality check. An otic capsule in an organism without a skull? We need to be looking for the semicircular canals of a labyrinth structure, not the hard tissue of a braincase.
4. The conodont apparatus is taken up in more detail here. According to the careful work of Purnell & Donoghue (1997), the conodont apparatus makes best sense if the affair is largely mounted on bilateral cartilage plates. Given the size of the apparatus and its obvious importance to the organism, these plates should be substantial. Where are they in the Crawling Eye? Similarly, Purnell (2001, 2001a) has shown that the conodont P-elements were capable precise occlusion. Again, this argues for substantial, possibly massive bilateral cartilages associated with the conodont apparatus in the head region. Where are they in the Crawling Eye model? Is it possible that the optic sclera and even the eye itself are partially preserved while these cartilages have completely disappeared?
5. What might be the purpose of such large, laterally placed eyes in what seems to have been -- by any other design criterion -- a benthic organism? Mud grubbers usually dispense with elaborate eyes relatively quickly.
6. Finally, while not every Paleozoic vertebrate has pineal and/or parapineal foramina, and they would not necessarily be preserved, one typically looks to find such structures in primitive chordates.
 None
of the problems above absolutely refute the Crawling Eye model of the conodont
head. We probably lack sufficient data to risk any kind of firm
conclusions at the present time. However we do assert that it is possible
to arrive at a more likely explanation for the observed structures -- one
that is more consistent with hard information from hard tissues and less
reminiscent of B-movies from the 1950's.
None
of the problems above absolutely refute the Crawling Eye model of the conodont
head. We probably lack sufficient data to risk any kind of firm
conclusions at the present time. However we do assert that it is possible
to arrive at a more likely explanation for the observed structures -- one
that is more consistent with hard information from hard tissues and less
reminiscent of B-movies from the 1950's.
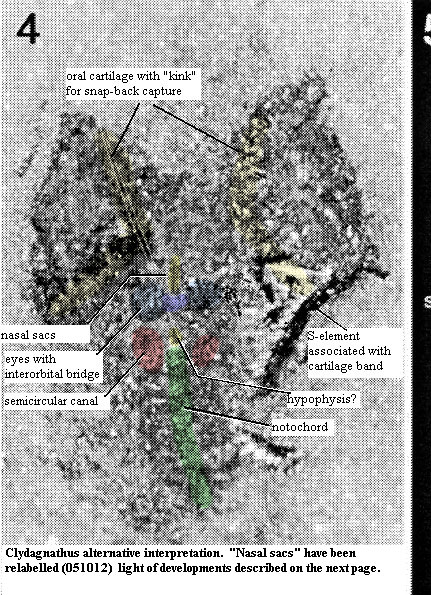
Consider, for example, the alternative model to the right. Other than the "eyes" of the Crawling Eye model, the conodont is revealed to be a very ordinary sort of vertebrate. It has two, primitive eyes which are closely paced, dorsal, and joined by an interorbital bridge. The eyes are closely associated with a nasohypophyseal foramen of some kind, just anterior to, and perhaps connected with hypophysis as in many other early craniates, such as the hagfish.
When suitably magnified, the semi-circular canals on the left side are unmistakable. There are no obvious gill pouches, but there is also no obvious need for them in an organism of this kind. Conodonts cannot have been filter-feeders. Purnell (2001). Thus, a pharyngeal filtering device is unlikely. A very small organism with the extreme elongation of a conodont would not require respiratory gills. Thus the posterior horizontal bands may be the arcualia of a true vertebrate, although they may also be respiratory structures if the organism habitually inhabited an oxygen-poor environment. The point here is that, whichever they are, they are not similar to, or serial homologues of, the semicircular canals.
So what are the "eyes" from the Crawling Eye? It is likely that they are precisely the missing oral cartilages of Purnell & Donoghue (1997). Purnell & Donoghue's structural studies of the ozarkodinid apparatus lead them to suggest that (a) the muscles controlling the apparatus must have operated through tendons running over a pulley - type structure and (b) that the apparatus may have "snapped" back into a retracted position from its everted state. Both of these requirements may be met if we suppose that the rims of the "eyes" are a curved or bent lateral oral cartilages and that the muscles were lateral to the cartilages. Thus the "eyeball" is revealed to be a mass of oral musculature and a complex tendon system. When the muscles contracted, tendons running around the lip of the cartilages from the lateral surface would pull the apparatus out and down, or in and up, exactly as envisioned by Purnell & Donoghue. Because the surface is curved, the distal ends of the S-elements would be spread like the ribs of a fan when everted. As the retractor muscles were contracted, these ends would snap inwards (medially) quite rapidly at first, then more slowly as the apparatus was drawn inwards.
ATW020727.
| Page Back | Unit Home | Page Top | Page Next |
checked ATW020917