| Palaeos: | Chondrichthyes | |
| The Vertebrates | Synechodontiformes |
| Page Back | Unit Home | Unit Dendrogram | Unit References | Taxon Index | Page Next |
| Unit Back | Vertebrates Home | Vertebrate Dendrograms | Vertebrate References | Glossary | Unit Next |
|
Abbreviated Dendrogram
Thelodonti |--Placodermi Eugnathostomata |--Chondrichthyes | |--Eugeneodontida | `--Chondrichthyes (Crown) | |--+--Cladoselachida | | `--Symmoriida | | |--Symmoriidae | | `--Holocephali | `--Elasmobranchii | |--Xenacanthida | `--Ctenacanthiformes | |--Ctenacanthidae | `--Euselachii | |--Hybodontiformes | `--+--Synechodontiformes | | |--Mucrovenator | | `--Palaeospinacidae | `--Neoselachii Teleostomi |
Contents
Overview |
The Synechodontiformes are Palaeospinax, Synechodus and that crowd. Our frank opinion is that, as normally considered, they are not a clade. We agree with numerous workers that the synechodontiforms are most likely a paraphyletic, and possibly polyphyletic, legion of basal Neoselachii who had the bad luck to become extinct before Owen and Agassiz reached drinking age. See, for example, Carroll (1988), deCarvalho (1996); Shirai (1996), and sources cited therein. Unfortunately, although this view is very widely held, opinion is not quite unanimous. In particular, Mikko Haaramo has shelved this group somewhere north of the ctenacanthiforms in one of his [former, as of 9/03] cladograms. Since Mikko is now a contributing editor to this site, he is technically part of the editorial "we." We rarely agree on any debatable point of phylogeny, and the result is a sort of evolutionary schizophrenia. Experience has taught us that Mikko has a distressing tendency to pick the winning side of the argument; and so, in a most rare, grudging and reluctant attempt to be a good sport, we will place this group of unmatched socks and single earrings in a middle ground -- a No-Man's-Land between the Mesozoic elsmobranchs and the living sharks and rays of the Neoselachii.
In the last analysis, this group may belong here. Although the modern literature refers to synechodontiforms as "neoselachian" sharks, this is actually a misnomer. The Neoselachii are a crown group: the last common ancestor of all living sharks and all of its descendants. There is no evidence that Synechodus, for example, belongs in this group. All we can say with confidence is that it is more closely related to living sharks than to the Hybodontiformes or the Ctenacanthidae. We cannot say that Synechodus is more closely related to some living sharks than to others. Maisey (1985) has argued, cogently, that Synechodus might be more closely related to the Galeomorphii than to the Squalea. However, even he is not prepared to commit himself on this issue. Synechodus, at least, is in the neoselachian stem group rather than in the Neoselachii itself.
The same does not apply to all members of the Synechodontiformes, as it has usually been conceived. It appears that Protospinax is actually well inside the Squalea -- the sister to the Hypnosqualea, a clade consisting of the rays, swordfishes, and Squatina. Maisey (1993). Accordingly, it has been transferred to that region and is not treated here.
 Palaeospinax:
Suppressed
Palaeospinax:
SuppressedThis is not the only, nor most famous, member of this group which has been phylogenetically expelled from the clade. Palaeospinax, the anchor of the Palaeospinacidae, is also gone. In this case, the sentence was more severe. Rather than banishment and exile among the rays, Palaeospinax has been suppressed altogether. It appears that Palaeospinax was based on essentially non-diagnostic material. Although good body-fossils were later referred to the genus, so were numerous, equally non-diagnostic teeth. The latter could not be differentiated from Synechodus in any systematic way. Worse, the body fossils were also indistinguishable from Synechodus. Accordingly, Palaeospinax has been declared a nomen dubium and all of this material referred to Synechodus. See Thies (1995). Indeed, there is some doubt as to whether even Paraorthacodus can be sufficiently distinguished from Synechodus. Thies (1991); Thies (1993). For the moment, however, Paraorthacodus has been granted a reprieve, at least on these pages.
In the early part of the last century, most workers thought that the Neoselachii descended from the hybodontiform sharks. However, it gradually became clear, due in large measure to the work of John Maisey in the 1980's, that this was probably not the case. The synechodontiforms share many characters with the Neoselachii which are quite unique and distinctive. That is, there are a large number of synapomorphies which characterize a clade consisting of Neoselachii + Synechodontiformes. Maisey (1984); see also, Carroll (1988), Maisey (1984a). The characters listed by Maisey may be said to involve the following functional groups:
 1.
Characters related to axial strength. The notochord is strongly reinforced
with bone. It is no longer "free," but segmented by vertebrae
and constricted. Calcification also occurs within the notochord,
between the internal and external elastic sheathes. DeCarvalho
(1996). The latter process is referred to as "primary
calcification" to distinguish it from the more familiar external or
"secondary" calcification. Where the skull
articulates with the notochord, it does so through distinct occipital condyles
which, like terrestrial vertebrates, are part of an occiput formed by
incorporating vertebral elements into the skull. In the stem group,
the condyles are formed from separate, identifiable
"hemicentra." DeCarvalho
(1996).
1.
Characters related to axial strength. The notochord is strongly reinforced
with bone. It is no longer "free," but segmented by vertebrae
and constricted. Calcification also occurs within the notochord,
between the internal and external elastic sheathes. DeCarvalho
(1996). The latter process is referred to as "primary
calcification" to distinguish it from the more familiar external or
"secondary" calcification. Where the skull
articulates with the notochord, it does so through distinct occipital condyles
which, like terrestrial vertebrates, are part of an occiput formed by
incorporating vertebral elements into the skull. In the stem group,
the condyles are formed from separate, identifiable
"hemicentra." DeCarvalho
(1996).
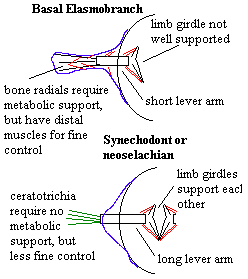
2. Concomitantly, the radial elements are somewhat lightened and simplified. Rather than growing, cyclomorial scales, neoselachians and synechodonts have simple, non-growing, placoid scales. In all likelihood, this is more expensive, as disposable items often are; but these scales are lighter, perhaps more hydrodynamic, and do not require as much dermal support or peripheral circulation, thus substantially reducing the amount of mass far from the axis. Fin spines may be present is some synechodontiforms, but the spines are generally simpler and smaller. Fin radials are generally reduced and replaced with lighter and more flexible ceratotrichia. The fin girdles are joined across the midline and the fin basals are more strongly attached to the girdles. What is going on here is that, once again, weight is being moved toward the center of the shark. With stronger basals and limb girdles, the muscles controlling the limbs be moved medially. This involves a loss of efficiency and fine control, offset by a gain in speed, since the axialized muscles need move only a small distance (albeit against a large force) to move the fins through a long lever arm.
 3.
Other changes occur in teeth and scales, which are functionally less
clear. The teeth have three enamel layers. The usual hybodont
pattern is two layers of enamel with parallel rods, referred to in the
literature as "PFE" or "single crystallite enameloid," one
more compact than the other. Synechodonts and neoselachians have three
layers: (a) they acquire an outer, shiny layer ("SLE") of highly
mineralized enamel with randomly oriented fibers, (b) a middle layer somewhat
similar to the hybodonts ("parallel bundled enameloid" or
"PBE") and (c) an inner layer with "haphazardly-oriented
fibers," "tangled bundled enameloid," "TBE", etc.
Unfortunately, both our perceptions and the nomenclature of shark enamel is in a
state of flux, largely due to the gradual and uneven adoption of scanning
electron microscopy as a diagnostic tool. See images at left from Cuny
et al. (2001).
3.
Other changes occur in teeth and scales, which are functionally less
clear. The teeth have three enamel layers. The usual hybodont
pattern is two layers of enamel with parallel rods, referred to in the
literature as "PFE" or "single crystallite enameloid," one
more compact than the other. Synechodonts and neoselachians have three
layers: (a) they acquire an outer, shiny layer ("SLE") of highly
mineralized enamel with randomly oriented fibers, (b) a middle layer somewhat
similar to the hybodonts ("parallel bundled enameloid" or
"PBE") and (c) an inner layer with "haphazardly-oriented
fibers," "tangled bundled enameloid," "TBE", etc.
Unfortunately, both our perceptions and the nomenclature of shark enamel is in a
state of flux, largely due to the gradual and uneven adoption of scanning
electron microscopy as a diagnostic tool. See images at left from Cuny
et al. (2001).
4. There are some changes in the jaw and braincase generally involving a looser postorbital articulation with the palatoquadrate. These changes should, however, be taken with considerable caution, since the whole business of loosening the jaw is reversed in most Squalea. However, there is no doubt that the braincase is very different from the hybodonts and ctenacanthids. Just for the sake of interest, we scored and ran the braincase of Synechodus, as described by Maisey (1985), on the character matrix developed by Coates & Sequiera (1998). While we would not dignify the results of this rough test by attempting to describe them in detail, it is clear that the braincase of Synechodus is very different indeed from the hybodont neurocranium, as well as strangely primitive.
Synechodontiformes: "Hybodus" minor, Sphenodus.
Range: Early Triassic to Paleocene (Eocene?)
Phylogeny: Euselachii:: Neoselachii + *: Mucrovenator + Palaeospinacidae.
Note: There is apparently a significant body of opinion that the synechodontiforms are within the crown group Neoselachii and are closer to the Squalea than to Galeomorphii. ATW020803.
Mucrovenator: Cuny et al. 2001. M. minimus Cuny et al. 2001.
Range: Middle Triassic (Anisian) of North America.
Phylogeny: Synechodontiformes: Palaeospinacidae + *.
Characters: [1] well-developed main cusp; main cusp recurved lingually, sometimes with sigmoid curve; main cusp with distal & mesial cutting edges extending entire height of cusp; 1-2 pairs of well-separated lateral cusplets; 1st pair never >50% height of main cusp; 2nd pair always smaller; cusps also with cutting edges; cutting edges not continuous between cusp/cusplets; ornamentation of parallel ridges ascending crown to apex; number of ridges variable (number correlates with height of cusplets); no furrow between root & crown; root very shallow; root extends lingually; root with long depression on labial side near base with row of foramina; lingual side of root shows large foramina (without depression?); SLE and PBE layers present in enamel; no TBE found [2]; in PBE there are thin radial enamel bundles which dominate in some areas of the tooth.
Notes: [1] All information from [c+01]. This is a tooth genus, and all characters are characters of teeth. [2] these are bundles of enamel. There are no fibers. See image above. The gross morphology of the teeth does not differ from the usual synechodont pattern. See, e.g., the images of Orthacodus teeth under Palaeospinacidae. [3] Cuny et al. (2001) are naturally troubled by the absence of a third, tangled, bundled layer. It may be, as they point out, that this layer was not preserved, or was lost in preparation of these very small, delicate teeth. This sounds like post hoc rationalization, but isn't. There seems to have been a good deal of variability in the recovery of this third layer in other fossils for reasons which are not clear. See, for example, DeCarvalho (1996: 54). However, it may also be that the TBE layer was a later development.
References: Cuny et al. (2001) [C+01]. ATW030329.
| Page Back | Unit Home | Glossary | Page Top | Page Next |
checked ATW090301