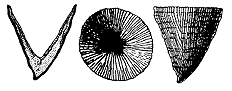
Lambeophyllum profundum (Conrad)
specimen about 2.5 cm long and in diameter
Blackriveran age (Harnagian)
New York
(reference: Moore, Lalicker & Fischer, Invertebrate Fossils, p.114)
| Late Ordovician Epoch | ||
| Paleozoic: Ordovician Period |
Katian Age (Late Caradoc to Middle Ashgill) |
| Sandbian | Middle Ordovician | Late Ordovician | ||
| Hirnantian | Llandovery | Time |
The Ordovician has been quite significantly revised in recent years. The traditional division of the Late Ordovician into the Caradoc and Ashgill has been replaced by a three-part division. The first two ages of the Late Ordovician are the Sandbian and Katian Ages. The Sandbian is equivalent to the Early Caradoc. The Katian is equivalent to the Late Caradoc and Early-Middle Ashgill. The Hirnantian, formerly the last stage of the Ashgill, is now the final age of the Ordovician. The base of the Katian is defined by reference to the first occurrence of the graptolite Diplacanthograptus caudatus, just below the base of the Guttenberg carbon-13 isotope excursion. The GSSP is located 4 m above the base of the Bigfort Chert, Black Knob Ridge section, 5 km northeast of Atoka, Oklahoma, USA.
When the ICS first set out to map the Ordovician in 1960, the task may have looked routine. The Ordovician System was about as old as stratigraphy itself, and the British system was used by nearly everyone, nearly everywhere in the world. As it turned out, this unanimity of historical practice masked conflicts in the data which could not be resolved. The foundation of the British Late Ordovician rested on a peculiarly British fauna which didn't play well with global graptolite zones. Consequently, almost everything had to be reworked. Chen et al. 2006). This is a good thing for us, because it has generated a great many high-quality stratigraphic studies within the last decade or so. Even so, local correlations have not been able to keep up with the changing names and biostratigraphy. Our usual desultory review of the literature has allowed us to construct only the following, and with many uncertainties at that:
| Global Timeslice Webby (2004) | North American Graptolite Zone Nardin (2007); Stott & Jin (2007) | ICS Stage | North America Stott & Jin (2007) | Britain Stott & Jin (2007) | Bohemia Nardin (2007), Ausich et al. (2002). | Baltic Paškevičius (2007) | Siberia Cocks & Torsvik (2007) |
| 6c | Normalograptus extraordinarius | Hirnantian | (Hirnantian) | Kosovian | Porkuni | Burian? | |
| 6b | Parorthograptus pacificus | (Gamachian) | (Rawtheyian) | Pirgu | Ketski | ||
| 6a | Dicellograptus complanatus & Diplograptus ornatus | (Cautleyian) | Vormsi? | Nirundian | |||
| 5d latest | (Richmondian)?? | Králodvorian (Avalonian Ashgill) | |||||
| 5d late | Ashgill (Pusgillian) | Nabala? | |||||
| 5d mid | Amplexograptus manitouliensis | (Maysvillian) | Dolbor? | ||||
| 5d early? | Geniculograptus pygmaeus | Cincinnatian (Edenian) | (Steffordian) | Rakvere? | |||
| 5c | Diplacanthograptus caudatus | Katian | (Trentonian?) | (Cheneyian-Steffordian) | Oandu | Baksan? | |
| 5b | Climacograptus bicornis | Sandbian | Mowhawkian (Black River?) | Caradoc (Burrellian) | Berounian | Keila | Chertov |
The Katian largely reflected the peak of the overall global warming trend of the Ordovician. However, there are some indications of a brief cold spell towards the middle of the Katian, perhaps coincident with the traditional Caradoc-Ashgill boundary. This is reflected in changes of sedimentation patterns in Baltica (Cocks & Torsvik, 2005) and a minor dip, or at least leveling off, in the global diversity profiles of many animal groups (Webby, 2004a). On the other hand, both may be explained by the merger of Avalonia, Baltica and Laurentia which began at this time. The closing of the northern Iapetus Ocean in the Katian would certainly have affected coastal currents in some fashion, and it clearly resulted in a sorting out of the endemic marine fauna around the three continents. Cocks & Torsvik (2005); Hints et al. 2007). The exact locations of Siberia and Perunica at this time are unclear, but they, too, seem to have been close enough to Baltica for some degree of faunal exchange to take place. Cocks & Torsvik 2002); Tychsen & Harper 2004); Cocks & Torsvik (2007).
What all this does not explain is the interesting pattern of radiolarian evolution. Radiolarian diversity peaked around the mid-Katian. The group seems to have begun its decline well-before the Hirnantian (Webby, 2004a), and this may suggest a generally less healthy open marine environment. A similar pattern is found in other environmentally-sensitive groups, such as sponges (Webby, 2004) and some brachiopods. Id.
Rugose Corals
Lambeophyllum profundum (Conrad)
specimen about 2.5 cm long and in diameter
Blackriveran age (Harnagian)
New York
(reference: Moore, Lalicker & Fischer, Invertebrate Fossils, p.114)
Order Orthida
Family Dalmanellidae

Paucicrura rogata (Sardeson)
shell about 7mm long and wide
Trenton Group
North America
more on Paucicrura
(at the Union College Geology Department site)
Order Strophomenida
Family Sowerbyellidae

Sowerbyella sericea (Sowerby)
Caradoc
England
shell about 1 cm wide
more on Swerbyella
(at the Union College Geology Department site)
|
|
|
Calyptaulax callicephalus, dalmanitid trilobite - family Pterygometopidae. Black River and Trenton Groups (Sandbian-Katian boundary), eastern and northeast Canada, and New York state. |
The Megalograptoids are, despite the name, eurypterids. Megalograptus is, in fact, the first apparently widespread genus of eurypterid, and its remains have been found in several Katian locations in eastern Laurentia from Quebec to Virginia. It is not certain that they actually evolved in Laurentia. The earliest known eurypterids are from the Sandbian of Wales. However, for most of their history, they remained largely in eastern Laurentia, Avalonia and western Baltica. Tetlie (2007).
Several substantial swimming trilobite groups evolved during the Late Ordovician, peaked in the Katian and became extinct shortly thereafter. Webby (2004a). Meanwhile, the characteristic Paleozoic arthropod fauna, including eurypterids, phyllocarids, and ostracodes, continued to expand. Id.
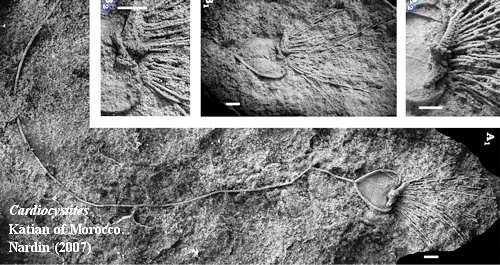 The Katian was an exuberant time for echinoderms. Almost all of the major echinoderm clades had Katian representatives: asteroids, crinoids, diploporans, eocrinoids, edrioasteroids, ophiuroids, rhombiferans, solutes, and stylophorans. Recent work on the eocrinoid Cardiocystites suggests that these primitively epibenthic forms evolved adaptations to lift themselves off the bottom, using long (10-20 cm), flexible stems to fix themselves to the bottom, while the theca floated above. This system presumably allowed these echinoderms to feed well above the bottom, even in high-energy environments, without being swept away. In fact, with this tether, a faster current would only increase the flow of food items available to the animal. The strategy seems to have been a success, as specimens of Cardiocystites are known from Britain, Gondwana (Morocco) and Perunica (Bohemia). Nardin (2007). This globalization is typical of Late Ordovician echinoderm groups. Ausich et al. (2002).
The Katian was an exuberant time for echinoderms. Almost all of the major echinoderm clades had Katian representatives: asteroids, crinoids, diploporans, eocrinoids, edrioasteroids, ophiuroids, rhombiferans, solutes, and stylophorans. Recent work on the eocrinoid Cardiocystites suggests that these primitively epibenthic forms evolved adaptations to lift themselves off the bottom, using long (10-20 cm), flexible stems to fix themselves to the bottom, while the theca floated above. This system presumably allowed these echinoderms to feed well above the bottom, even in high-energy environments, without being swept away. In fact, with this tether, a faster current would only increase the flow of food items available to the animal. The strategy seems to have been a success, as specimens of Cardiocystites are known from Britain, Gondwana (Morocco) and Perunica (Bohemia). Nardin (2007). This globalization is typical of Late Ordovician echinoderm groups. Ausich et al. (2002).
As discussed in at various other points, a relatively complete set of primitive vertebrates had also appeared by the Katian. Any Ordovician vertebrate remains are still rare, but thelodonts, in particular, become considerably more common in the Katian -- specifically in Timan-Pechora and other terranes of present-day Asia. The Katian (or perhaps Sandbian) also produced the last major group of early Paleozoic vertebrates, the Acanthodii, a group of fish close to our own ancestry. Sansom et al. 2001). Since acanthodians and sharks are both known by the Katian, we can be relatively certain that jaws had evolved by this time. This useful addition to the vertebrate repertoire seems to have taken a little while to perfect, since the great radiation of gnathostome fishes was still more then 20 My in the future.
ATW080405
checked ATW060130
